





Puesta a punto del modelo de HP en ratones
Durante el desarrollo de las actividades de investigación fueron necesarias modificaciones, dentro del PQE, para lograr el mayor éxito posible durante la cirugía de los roedores. Particularmente, el principal obstáculo que impedía una mayor tasa de éxito de la cirugía fue la anestesia administrada. Como se mencionó previamente, se utilizó una mezcla de ketamina/xilacina por administración intraperitoneal, iniciando los estudios con una dosis de los fármacos dentro del rango propuesto en el PQE que se llevó adelante para la realización de las hepatectomías (Nevzorova Y, Tolba R, Trautwein C, & Liedtke C, 2015). Sin embargo, la dosificación debió ser modificada a lo largo de la investigación debido a que resultaba excesiva para ambas cepas de ratones y en mayor medida para la cepa knockout para Kir6.2 (Kir-/-). La intoxicación provocaba que los roedores murieran como consecuencia de una depresión del sistema nervioso central inducida por la anestesia, con posterior falla del sistema respiratorio y paro cardiovascular.
Se comenzaron las cirugías utilizando una solución de ketamina/xilacina de 100 y 3 mg/kg PC, respectivamente. Utilizando esta dosis de anestésicos, se realizó la HP a 4 ratones WT y 3 ratones Kir-/-. De los ratones WT hepatectomizados, 2 ratones murieron; mientras que de los 3 ratones Kir-/- ninguno sobrevivió a la cirugía. Se decidió disminuir la dosis al 50% para la cepa Kir-/- (ketamina/xilacina 50 y 1,5 mg/kg PC, respectivamente), y se mantuvo la dosis inicial para los ratones WT. Se obtuvieron los siguientes resultados: de los 18 ratones WT hepatectomizados, 7 sobrevivieron al procedimiento, dando una tasa de mortalidad del 61%. A su vez, de las 15 cirugías realizadas a ratones de la cepa Kir-/- al 50% de la dosis, se obtuvo una tasa de mortalidad del 53% (8 ratones murieron). A esta dosis, hacia el final de la cirugía los efectos anestésicos iban desapareciendo, por lo que se administró éter dietílico como anestésico inhalatorio, en cada caso según necesidad. En una última modificación de la dosis de anestésicos, se procedió a administrar el 75% de la dosis (ketamina/xilacina 75 y 2,25 mg/kg PC, respectivamente) a ambos grupos de ratones, aumentando la dosis en Kir-/- y disminuyendo la dosis en cepa WT con respecto a la primera modificación realizada. Se realizaron un total de 18 cirugías en ratones WT, obteniendo una tasa de mortalidad del 26% (5 ratones murieron); mientras que en ratones de la cepa Kir-/- se realizaron 15 HP, y solo un ratón murió, resultando en una tasa de mortalidad del 6% para esta última modificación de la dosis de anestesia.
La ausencia de Kir6.2 no modifica el patrón de actividad de las transaminasas hepáticas durante la regeneración hepática posterior a una HP
Se analizaron en suero el perfil de las enzimas ALT y AST, marcadores de daño hepático. La medición fue realizada a diferentes tiempos: 0, 1, 2, 3, 48, 96 y 168 h post HP. Tras el análisis de los datos se observó el perfil típico de aumento de estas enzimas posterior a una HP, en relación directa a la injuria hepática que se produce durante la resección. En ambas cepas se constató un leve aumento en ALT y AST a partir de las 3h post HP y un pico en la actividad de dichas enzimas a las 48 h, normalizando hacia tiempos más prolongados (figura 3).
Estos resultados fueron coincidentes con el aumento de transaminasas que se produce posterior a una HP (Mueller A-K, Senninger N, & Vowinkel T, 2017). En consecuencia, podemos afirmar que el patrón de actividad enzimática en suero tras la lesión hepática generada por la cirugía se encontró conservado tanto en la cepa WT como en la cepa Kir-/-. No se observa diferencia significativa entre los grupos estudiados a los diferentes tiempos, por lo que se puede inferir que la ausencia de Kir6.2 no produce una alteración en el perfil de cambio de estas enzimas posterior a la hepatectomía.
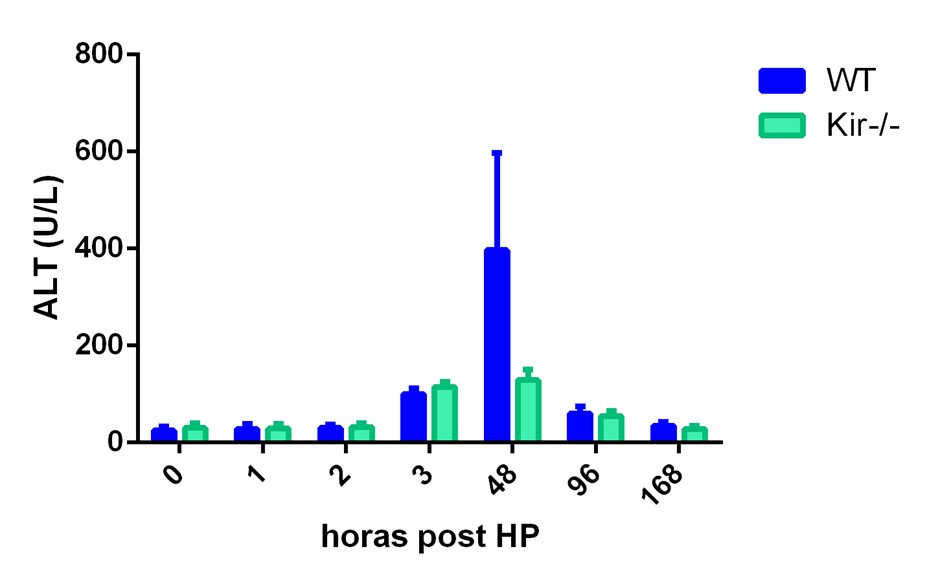
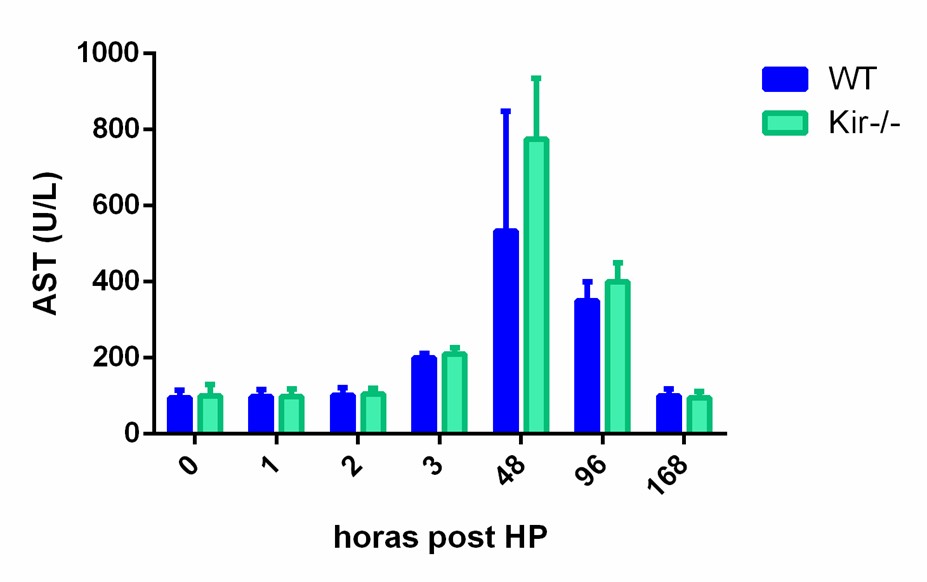
FIGURA 3. Perfil de actividad enzimática de las transaminasas AST y ALT en suero de ratones WT y Kir-/- a diferentes tiempos post HP. Los resultados se expresan como media ± SEM (n= 3-4 ratones por condición).
La ausencia de Kir6.2 reduce el índice peso hígado/peso corporal durante la regeneración hepática posterior a una HP
La figura 4 muestra el índice PH/PC a 0, 1, 2, 3, 48, 96 y 168 h posteriores a una HP, en ratones controles y carentes de la proteína Kir6.2. Como se observa en la figura, a todos los tiempos post HP estudiados hay una diferencia significativa en el índice entre los animales WT y los Kir-/-, estos últimos mostrando un índice considerablemente menor.
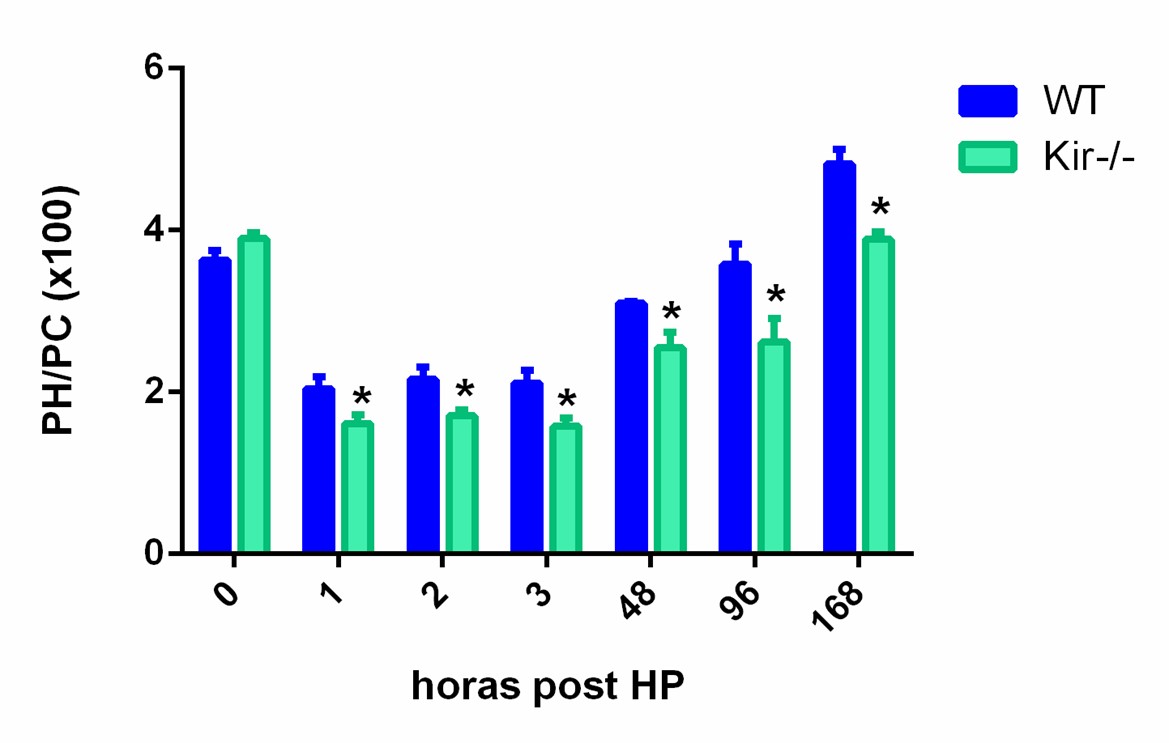
FIGURA 4. Índice PH/PC (x100) correspondiente a los tiempos 0, 1, 2, 3, 48, 96 y 168 h post HP para ratones de la cepa C57/BL6 (WT) y Kir6.2-/- (Kir-/-). Los resultados se expresan como media ± SEM (n= 3-4 ratones por condición); *p< 0,05 vs. su correspondiente WT para cada tiempo.
La ausencia de Kir6.2 no afecta la histoarquitectura hepática durante la regeneración hepática posterior a una HP
Los cortes histológicos de hígados de ratones WT y Kir-/- a los tiempos 0, 48 y 168 h post HP fueron teñidos con hematoxilina y eosina y analizados por un especialista (Bioq. Gerardo Pisani, Servicio de Histotecnología, Facultad de Ciencias Bioquímicas y Farmacéuticas, UNR). Se eligieron los mencionados tiempos para analizar la histoarquitectura hepática al momento de la cirugía (0 h), al tiempo donde se produce un pico de proliferación de los hepatocitos (48 h) y al final del proceso regenerativo (168 h).
La figura 5 muestra imágenes representativas. La observación de los preparados histológicos analizados mostró que los hígados de los ratones Kir-/- presentaban una histoarquitectura conservada, sin signos de inflamación ni necrosis, al igual que en los animales WT. Por otra parte, en los cortes histológicos a las 48 h post HP se observó un marcado número de mitosis, indicativo del estado proliferativo de los hepatocitos.
Cortes histológicos de hígados de ratones de la cepa C57/BL6 (WT) y Kir6.2-/- (Kir-/-) teñidos con hematoxilina y eosina
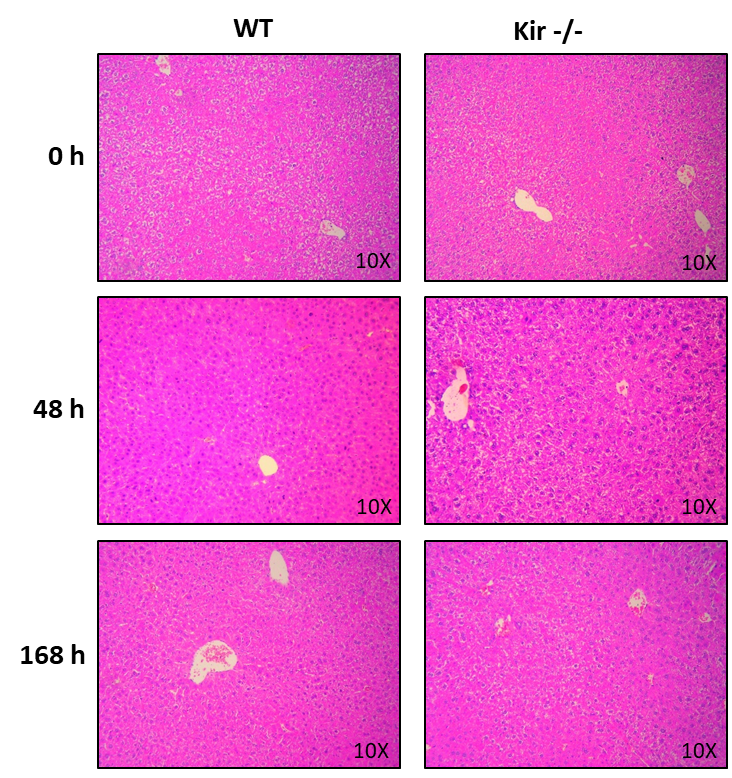
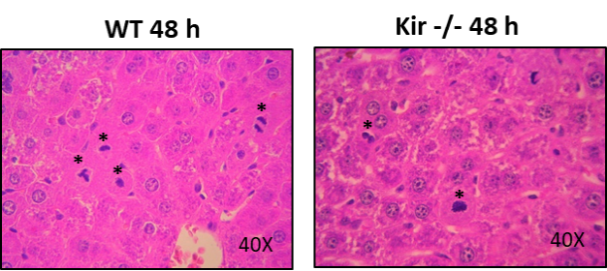
FIGURA 5. En los primeros tres paneles se muestran imágenes representativas a 0, 48 y 168 h post HP. Aumento: 10X. En el panel de abajo se muestran imágenes de hepatocitos en mitosis (marcados con *) a las 48 h post HP. Aumento: 40X.
La ausencia de Kir6.2 reduce el estado proliferativo de los hepatocitos durante la regeneración hepática posterior a una HP
Dado que el índice PH/PC se mostró significativamente disminuido en los animales Kir-/-, lo cual indica una disminución en la recuperación de la masa hepática posterior a una HP, se decidió estudiar el estado proliferativo de los hepatocitos en el hígado remanente. Para ello, se realizó la tinción inmunohistoquímica de PCNA a 0 h y a las 48 h post HP, tiempo en el que existe un pico inicial de síntesis de ADN posterior a una HP en ratón. PCNA es una proteína auxiliar de la ADN polimerasa delta, que juega un papel crítico en la iniciación de la proliferación celular. Aunque PCNA se correlacionó con la fase S del ciclo celular hoy se sabe que, si bien su pico se encuentra en dicha fase, en realidad se expresa durante todo el ciclo. Es expresado, por tanto, por todas las células proliferantes, pero no por las células quiescentes o en reposo, es decir en fase G0. Este marcador, además, permite diferenciar la fase del ciclo celular en la cual se encuentra la célula, ya que cada fase presenta un patrón de tinción especifico (Foley JF, Dietrich DR, Swenberg JA, & Maronpot RR, 1991).
En la figura 6 se muestran imágenes de la detección inmunohistoquímica de PCNA en hígados de ratones de ambas cepas, WT y Kir-/-. Como era de esperar, el número de hepatocitos PCNA positivos al momento de la hepatectomía (0 h) es muy reducido, mientras que aumenta marcadamente a las 48 h, siendo este aumento mayor en el grupo WT en comparación con el grupo Kir-/-. Como se mencionó previamente, la proteína PCNA es expresada por los hepatocitos en estado proliferativo, y no en G0. Se podría afirmar, por lo tanto, que los hepatocitos en los animales Kir-/- están en mayor proporción en estado quiescente y no entraron en proliferación. Cabe destacar que tampoco se apreció una diferencia significativa entre ambos tiempos de la misma cepa. Estos resultados ponen de manifiesto que la proliferación de los hepatocitos se ve afectada por la ausencia de canales de K+ sensibles a ATP.
Para obtener una medida cuantitativa del estado proliferativo disminuido en los hígados regenerantes de los ratones Kir-/-, analizamos por Western Blot los niveles proteicos de PCNA y de ciclina D1. La ciclina D1 es una proteína clave para el ingreso de las células al ciclo celular y su posterior progresión a través de él (Albrecht JH, Poon RY, Ahonen CL, Rieland BM, Deng C, & Crary GS, 1998).
Inmunohistoquímica para PCNA
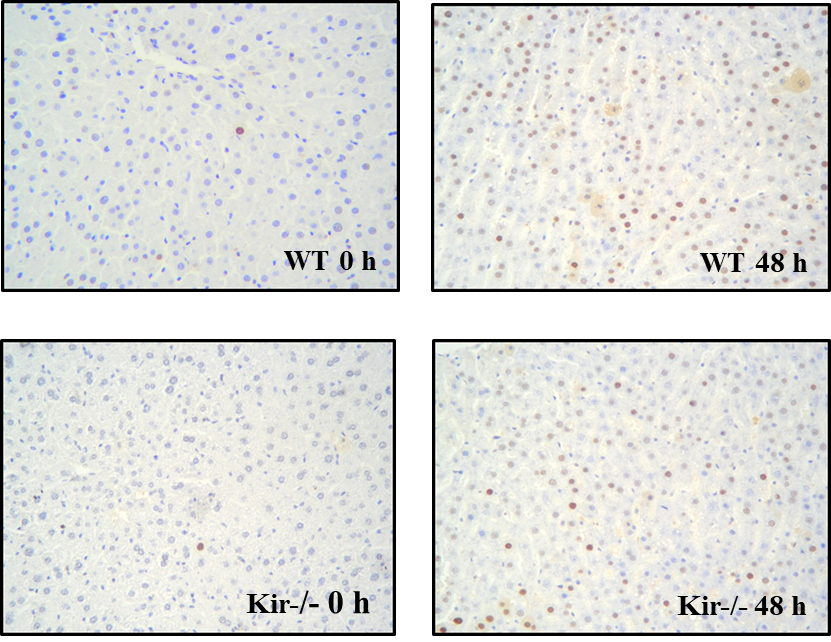
FIGURA 6. Imágenes representativas obtenidas mediante microscopía óptica a partir de muestras de hígado de ratones de la cepa C57/BL6 (WT) y Kir6.2-/- (Kir-/-) a las 0 y 48 h post HP. Aumento: 20X.
Estudios por Western Blot de la expresión proteica de PCNA y Ciclina D1
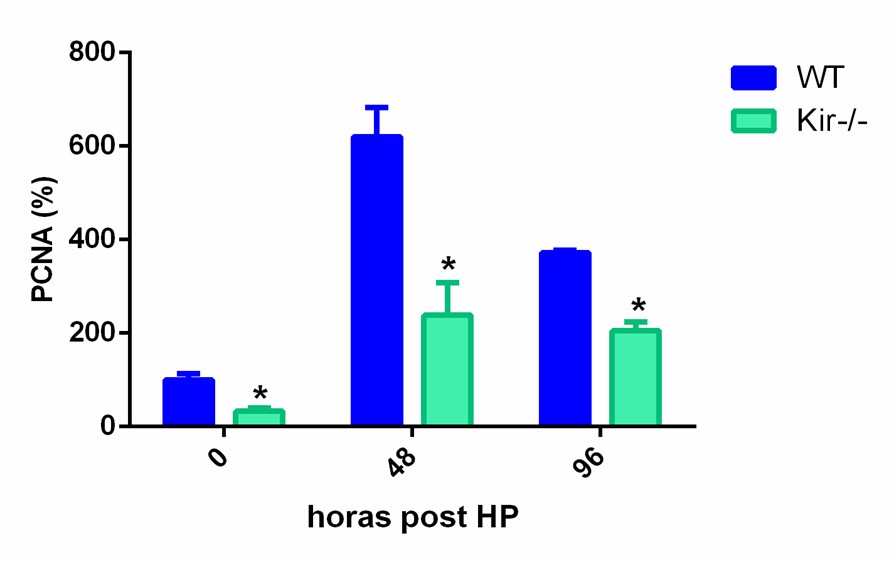
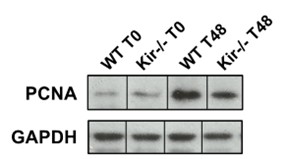
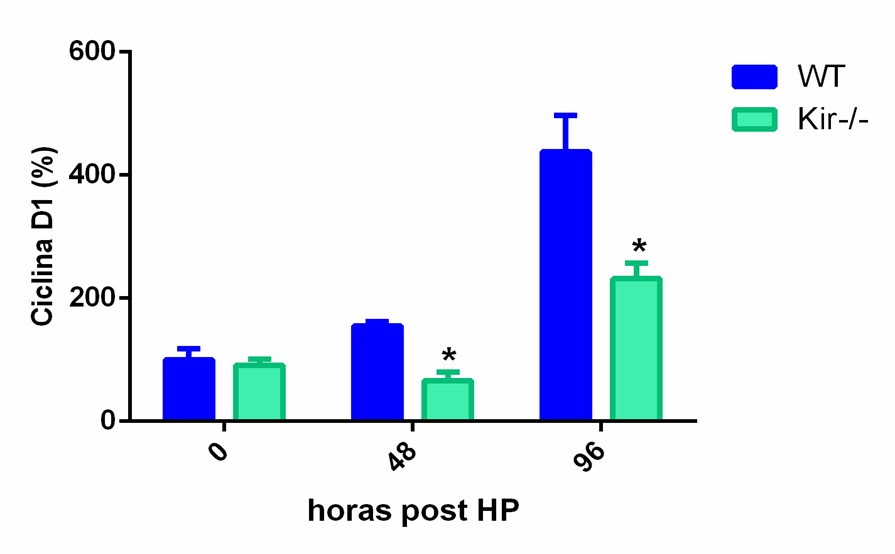
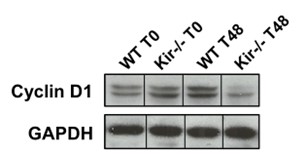
FIGURA 7. Estudios por Western Blot de la expresión proteica de PCNA y Ciclina D1 en lisados totales de muestras de hígado de ratones de la cepa C57/BL6 (WT) y Kir6.2-/- (Kir-/-) a las 0, 48 y 96 h post HP. A la izquierda se muestran las cuantificaciones, expresadas como porcentaje de los niveles del grupo WT a tiempo cero. Las barras representan la media ± SEM (n = 4 por condición). *p< 0,05 vs. su correspondiente WT para cada tiempo. A la derecha se muestran imágenes representativas de los revelados. La proteína GAPDH se utilizó como control de carga.
La ausencia de Kir6.2 aumenta la apoptosis durante la regeneración hepática posterior a una HP
Se ha descripto que en la regeneración hepática post HP, al final de la síntesis de ADN se produce una pequeña ola de apoptosis de los hepatocitos como un mecanismo para corregir el disparo de una respuesta regenerativa excesiva (Sakamoto T, Liu Z, Murase N, Ezure T, Yokomuro S, Poli V, Demetris AJ, 1999). Se decidió estudiar el nivel de apoptosis en los grupos WT y Kir-/- a través de la detección de proteínas de la familia Bcl-2, una familia clave que participa en la muerte apoptótica. Las proteínas de esta familia son reguladores de los cambios estructurales en la membrana mitocondrial que conducen a la apoptosis y hay miembros de la familia, como Bax, con función proapoptótica, es decir favorecen la muerte celular programada, mientras que otros miembros como Bcl-2 y Bcl-xL ejercen acciones antiapoptóticas (Edlich J, 2018). En el presente trabajo, se determinaron por Western Blot los niveles proteicos de Bax, Bcl-2 y Bcl-xL en lisados totales de hígado de ratones WT y Kir-/-, a las 0, 48 y 96 h post HP.
En cuanto a las proteínas antiapoptóticas, se determinó Bcl-xL a las 48 h post HP, mientras que Bcl-2 se midió a las 96 h post cirugía. Como era de esperar, en los animales control, los niveles de estas proteínas disminuyeron a estos dos tiempos con respecto a sus niveles basales. Cuando se compararon los niveles las proteínas en el grupo Kir-/- con respecto al grupo WT, ninguna de las dos proteínas antiapoptóticas mostró diferencias significativas entre las cepas. Bcl-xL mostró una tendencia a estar disminuida fundamentalmente al momento de la HP en los ratones Kir-/-, mientras que Bcl-2 mostró una tendencia a aumentar a las 96 h en la misma cepa (figura 8).
Estudios por Western Blot de la expresión proteica de las proteínas antiapoptóticas y proapoptóticas
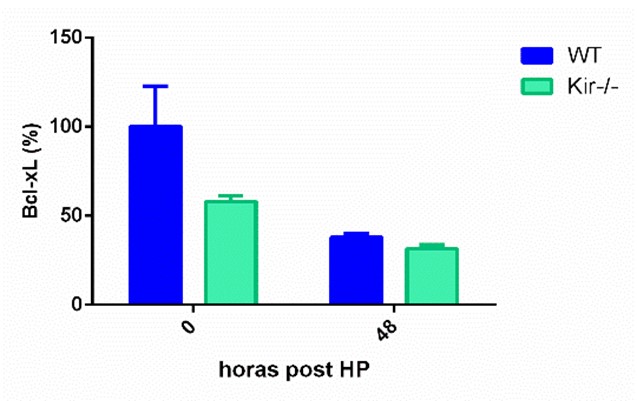
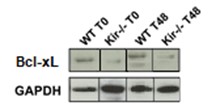
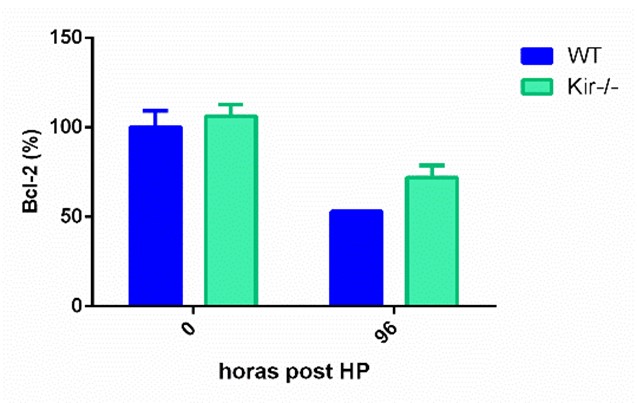
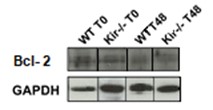
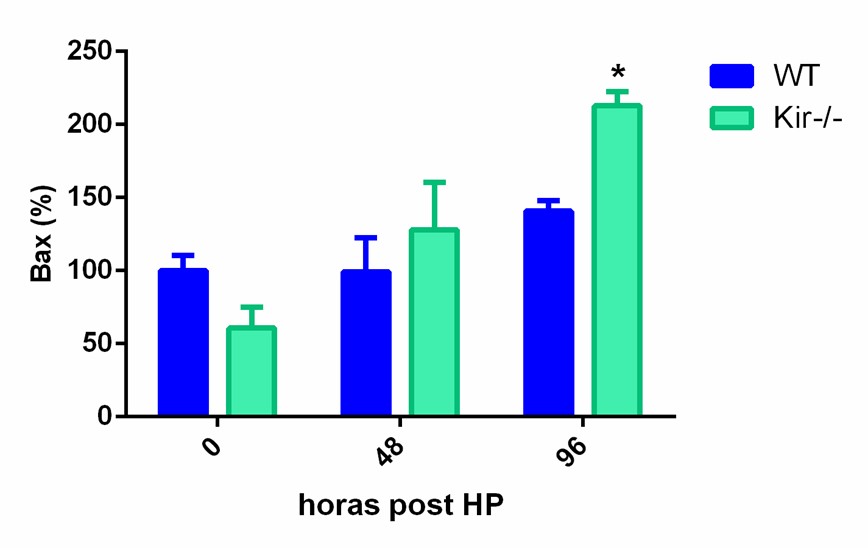
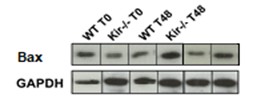
FIGURA 8. Estudios por Western Blot de la expresión proteica de las proteínas antiapoptóticas Bcl-xL (panel superior) y Bcl-2 (panel central) y de la proteína proapotótica Bax (panel inferior) en lisados totales de muestras de hígado de ratones de la cepa C57/BL6 (WT) y Kir6.2-/- (Kir-/-) a las 0, 48 y 96 h post HP. A la izquierda se muestran las cuantificaciones, expresadas como porcentaje de los niveles del grupo WT a tiempo cero. Las barras representan la media ± SEM (n = 3 por condición). *p< 0,05 vs. su correspondiente WT para cada tiempo. A la derecha se muestran imágenes representativas de los revelados. La proteína GAPDH se utilizó como control de carga.
La determinación de Bax arrojó resultados más interesantes, ya que se observó un patrón de aumento de dicha proteína que comenzó a las 48 h en la cepa Kir6.2, y se hizo significativamente mayor a las 96 h post cirugía en comparación con los ratones WT.
La prevalencia relativa de la proteína Bax y Bcl-2/Bcl-xL es un factor crítico que influencia el destino celular, promoviendo la supervivencia o la muerte, cuyo resultado depende en gran medida de la relación Bax/Bcl-2 o Bax/Bcl-xL. Así, la ruta de la apoptosis puede ser activada bajo condiciones en las que la expresión de la proteína Bax está elevada y/o la expresión de las proteínas Bcl-2/Bcl-xL está disminuida. En este estudio, se procedió a calcular el índice apoptótico Bax/Bcl-2 y Bax/Bcl-xL, y se observó que el mismo, a tiempo cero, arrojó valores similares en ambas cepas. A las 48 h, el índice apoptótico aumentó en ambas cepas en comparación con su valor a tiempo cero; sin embargo, resultó ser significativamente mayor en los hígados de los ratones Kir-/- con respecto a los WT. Por otro lado, el índice apoptótico a las 96 h continuó elevado en ambas cepas con respecto a sus niveles iniciales y, además, los animales carentes de Kir6.2, mostraron una tendencia no significativa a tener un índice apoptótico mayor que el de los animales WT a ese tiempo (figura 9).
Índice apoptótico
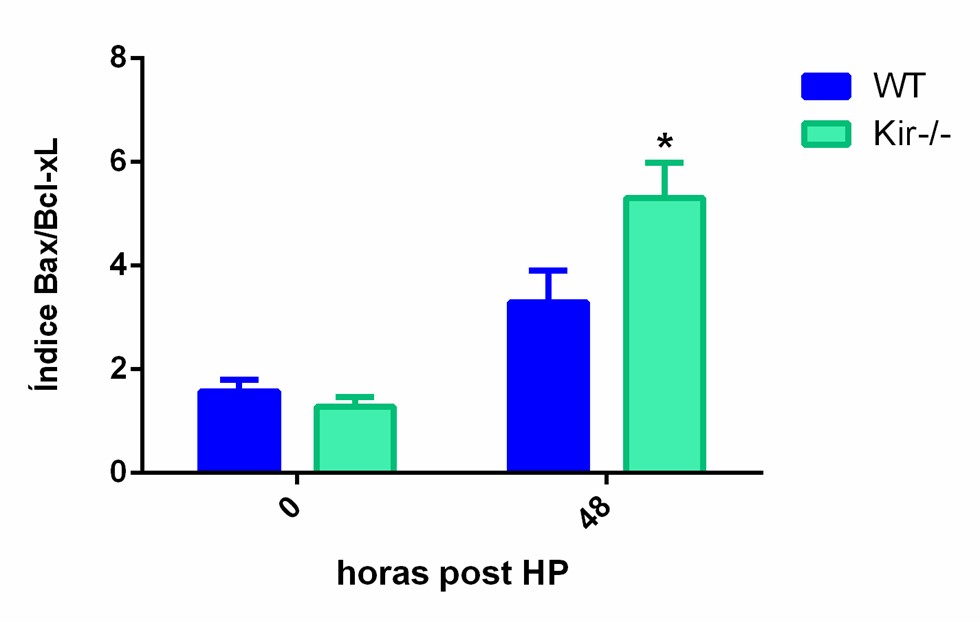
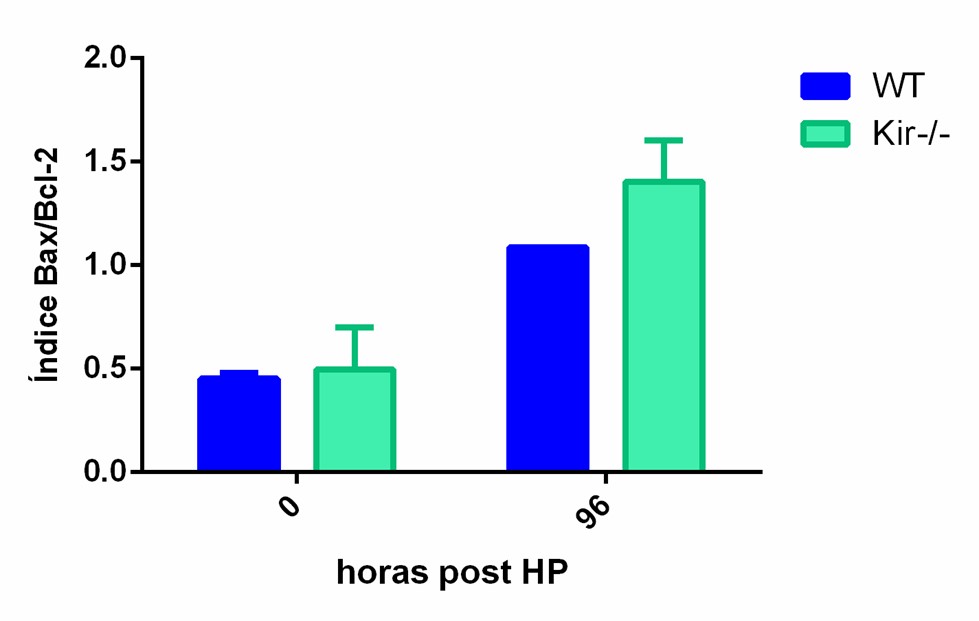
FIGURA 9. Índice apoptótico determinado en muestras de hígado de ratones de la cepa C57/BL6 (WT) y Kir6.2-/- (Kir -/-) a las 0, 48 y 96 h post HP. El índice apoptótico fue calculado como el cociente entre la proteína proapoptotica Bax y las proteínas antiapoptoticas Bcl-2/Bcl-xL. Las barras representan la media ± SEM (n=3 por condición) *p<0.05 vs su correspondiente WT para cada tiempo.

