





Un caso de estudio
Música porque sí, música vana,
como la vana música del grillo,
mi corazón eglógico y sencillo
se ha despertado grillo esta mañana.“El Grillo”, Conrado Nale Roxlo
El objetivo principal de esta tesis es ofrecer una elucidación del lenguaje funcional utilizado en la neuroetología. Como explicité en la introducción a este trabajo, mi estrategia para abordar dicha tarea reside en tomar una investigación específica de esta área y explicitar el uso del lenguaje funcional al que se apela en ella, para luego evaluar la adecuación de los enfoques presentados en el capítulo anterior a este caso de estudio. En virtud de dicho objetivo, a lo largo de este capítulo me enfocaré en presentar, de manera informal, el caso de estudio que servirá de fundamento para la evaluación de las elucidaciones precedentes, la cual será abordada en el capítulo siguiente.
El caso sobre el cual me basaré es la investigación neuroetológica concerniente a las bases neurales de la comunicación acústica del grillo de campo (Gryllus campestris y Gryllus bimaculatus). Los motivos para esta elección pueden resumirse en dos. En primer lugar, en el ámbito de la neuroetología, suele considerarse que los animales invertebrados en general, y los insectos en particular, constituyen buenos “sistemas modelos” para la investigación neuroetológica, en tanto exhiben comportamientos relativamente simples, robustos y accesibles, a la vez que poseen sistemas nerviosos constituidos por células relativamente grandes e identificables por los métodos neurobiológicos actuales. Por razones similares, este caso constituye un buen “sistema modelo” para la investigación filosófica, en tanto su relativa simplicidad lo convierte en un caso permeable y accesible para la búsqueda de patrones explicativos y regularidades en el uso de conceptos teóricos. En segundo lugar, las investigaciones biológicas dedicadas a la comunicación en los insectos, en especial en los grillos (Orthoptera: Gryllidae), langostas (Orthoptera: Acrididae), saltamontes (Orthoptera: Tettigoniidae) y cigarras (Hemiptera: Cicadidae), están entre aquellas que han logrado mayor desarrollo en el área, encontrándose entre las más completas no sólo en lo que concierne a la indagación sobre las bases neurales, sino en los diversos aspectos que abarca el estudio biológico del comportamiento. En virtud de ello, considero que abordar el lenguaje funcional de la investigación concerniente a la comunicación acústica en el grillo de campo puede ofrecernos claves valiosas para comprender el lenguaje funcional involucrado en la neuroetología en general, y cómo este lenguaje relaciona a la neuroetología con otras áreas dedicadas al estudio biológico del comportamiento.
El orden en el que abordaré la cuestión que nos ocupa es el siguiente. En el próximo apartado ofreceré una introducción general al estudio de la comunicación en los insectos y al caso de estudio en particular. En la sección 4.2 trataré la cuestión de la “función” del canto de llamada (cf. sección 4.1). En la sección 4.3 me explayaré sobre la indagación científica en torno a las bases neurales de la fonotaxis (cf. sección 4.1). Finalmente, en la sección 4.4 ofreceré un resumen de las cuestiones tratadas en este capítulo que me permitirán abordar, en el capítulo siguiente, la elucidación propiamente dicha del lenguaje funcional utilizado por los neuroetólogos.
Antes de comenzar, es importante señalar que la presentación del caso supondrá la utilización de conceptos metateóricos (como estructura, función, efecto) que aquí trataré de manera presistemática. Esto es, aquí enfatizaré la aparición de dichos conceptos y su rol en las diferentes instancias de la investigación de la comunicación acústica del grillo de campo, y utilizaré estos conceptos de manera cercana a los usos propios de los autores. Luego, en el capítulo siguiente, me avocaré a reflexionar sobre estos usos utilizando las teorías filosóficas presentadas precedentemente, utilizando un lenguaje metateórico más técnico.
4.1. Introducción al caso: el estudio de la comunicación en los insectos
Los insectos son la clase de organismos con mayor riqueza de especies en el planeta. Divididos en 30 órdenes, representan aproximadamente el 70 por ciento de las especies de la Tierra, y existen en una gran variedad de formas y ecosistemas (Chapman, 2013). Tal vez uno de los aspectos más llamativos de su comportamiento sean los sonidos que producen, especialmente durante las noches de verano, insectos como los grillos, las cigarras y los saltamontes. Tales sonidos son referidos a menudo como “cantos”, posiblemente debido a la estructura rítmica que los caracteriza (Alexander, 1960), y fueron notados por los estudiosos de la naturaleza desde la antigüedad (Bodson, 1983). La idea de que el comportamiento acústico de los insectos es un fenómeno asociado a su reproducción parece ser antigua. Ya Aristóteles, en su Historia Sobre los Animales, afirma que la “música” que producen los insectos, y los animales en general, está relacionada de alguna manera con su forma de reproducirse:
Unos animales emiten sonidos, otros son mudos y otros están dotados de voz: entre estos últimos, unos tienen un lenguaje articulado y otros inarticulado. Y unos son charlatanes y otros taciturnos; unos cantarines y otros no aficionados al canto. Pero común a todos es cantar o cacarear sobre todo en la época de apareamiento (Aristóteles, Historia de los Animales, libro I 6, 488a34).
Afirmaciones similares pueden hallarse, más de dos milenios después, en el propio Darwin:
Todos los observadores están de acuerdo en que los sonidos sirven para llamar o excitar a las hembras mudas. Pero ha sido informado que el macho de la langosta migratoria de Rusia (una de las Acridiidae), mientras está emparejado con una hembra, estridula de enojo o celos cuando es enfrentado por otro macho (Darwin, 1871, p. 352).
A pesar de estas antiguas observaciones, fue principalmente durante los últimos 60 años que las investigaciones sobre la producción, la percepción y el rol de tales sonidos en el comportamiento de los insectos lograron un desarrollo significativo y sistemático. De manera general, podría decirse que los estudios morfológicos dedicados a los órganos de producción y percepción de sonidos precedieron en buena medida a aquellos dedicados su rol en la conducta. Así, por ejemplo, los primeros estudios anatómicos detallados de los órganos auditivos de distintos grupos de insectos datan de principios del siglo XX[1] y, a mediados de siglo, ya se contaba con dos importantes estudios comprehensivos acerca del tema: “Hearing in Insects” (Pumphrey, 1940) e “Insect Sounds” (Haskell, 1961). Sin embargo, hasta ese momento, los estudios experimentales dedicados a la conducta habían sido escasos. Entre los pocos casos de experimentación previos a la primera mitad del siglo XX figura la demostración de Regen (1913) de que las hembras de grillo son atraídas por los cantos de un macho transmitidos a través de un teléfono. De este modo, se mostraba por primera vez la respuesta de aproximación de una hembra, o fonotaxis, ante el canto de llamada de un macho. El desarrollo de nuevas técnicas de grabación, análisis, manipulación y reproducción de sonido luego de la Segunda Guerra posibilitó e impulsó nuevos estudios en este área (Drosopoulos, 2006; Hedwig, 2014) y, partir de la década de los 60, el número de publicaciones acerca del tema incrementó rápidamente. Buena parte de la investigación experimental se ocupó del orden de los ortópteros, concentrándose en las respuestas de las hembras ante los cantos de los machos (Falls, 1992). En este contexto surge la investigación sobre la comunicación acústica del grillo de campo europeo (Orthoptera: Gryllus campestris y Gryllus bimaculatus). Como veremos, buena parte de los estudios experimentales realizados involucraron la reproducción de cantos naturales o modificadas digitalmente desde altavoces ubicados en diferentes direcciones y la utilización de diversos aparatos diseñados para medir la respuesta de las hembras. Tales estudios permitieron establecer qué propiedades de los cantos de llamada de los grillos son relevantes para provocar la respuesta fonotáctica de la hembra (su “estímulo señal”) y, junto con la utilización de técnicas de trazado neuronal y registro intracelular, permitieron indagar las redes neurales involucradas en dicha respuesta (su “mecanismo desencadenador”).
4.2. El comportamiento
En la mayoría de las especies de gríllidos (Gryllidae), los machos emiten su canto de llamada mediante un mecanismo de estridulación ubicado en sus alas delanteras. Durante la estridulación, los machos levantan las alas y, mediante movimientos de apertura y cierre, frotan la una sobre la otra a un ritmo constante y específico (Drosopoulos y Claridge, 2006). De esta manera, el rascador, una protuberancia de cutícula del borde interior del ala, raspa la lima, una fila de dientes espaciados regularmente ubicada en el envés del ala.
El sonido producido de esta manera posee una estructura consistente en una serie de pulsos, a menudo denominados “sílabas”. Cada cierre completo de las alas produce una única sílaba, y los cantos de llamada de los grillos de campo consisten típicamente en un tren de cuatro sílabas ─denominado “chirrido”─ seguido de un periodo de silencio (Zupanc, 2010). El canto producido de este modo posee la siguiente estructura:

Figura 2: Estructura del canto de llamada del grillo de campo (Gryllus bimaculatus). Pulse duration – Duración de la sílaba. Pulse pause – Duración del silencio entre sílabas. Chirp duration – Duración del chirrido. Chirp pause – Duración del silencio entre chirridos (Meckenhäuser G, Hennig RM, Nawrot MP, 2013).
Los machos comienzan a realizar sus llamadas varios días después de su muda adulta, cuando un espermatóforo está presente en su tracto genital (Zuk y Simmons, 1997). Los machos de las especies Gryllus campestris y bimaculatus cantan desde sus madrigueras, y las hembras sexualmente receptivas son atraídas por el canto y se aproximan a los machos, esto es, responden con el comportamiento denominado “fonotaxis” (Zupanc, 2010). Luego de que los sexos entran en contacto, los machos a menudo producen una “canto de cortejo”, de menor intensidad que el canto de llamada (Alexander, 1957). La copulación consiste en la hembra montando al macho, seguido de la transferencia de un único espermatóforo desde el macho hacia los genitales externos de la hembra (Gullan y Cranston, 2014). Inmediatamente después de la copulación, las hembras pueden remover o comer el espermatóforo y los machos pueden intentar prevenirlas de hacer eso realizando una serie de comportamientos estereotípicos a menudo denominados “guardia postcopulatoria” (Alexander, 1967).
4.3. La función del canto de llamada
La cuestión acerca de la “función” de los cantos producidos por los grillos, y por los insectos en general, fue objeto de especulaciones desde, como vimos, la antigüedad griega. Sin embargo, el estudio sistemático y comparado de tales sonidos dio lugar a ciertos estudios que lograron establecer con mayor precisión los diferentes roles o funciones que los cantos cumplen dentro de los patrones de comportamiento de estos animales.
Entre los estudios comprehensivos dedicados a establecer las funciones que cumplen los cantos de los ortópteros en general, y de los grillos en particular, se encuentran aquellos de Richard Alexander[2] (1957, 1960, 1961, 1962, 1967, entre otros). Tales estudios son ilustrativos del modo en el que los biólogos atribuyen “funciones” a los comportamientos y, como veremos más adelante, dicha atribución funcional es y fue fundamental para indagar las bases neurales de la fonotaxis.
Alexander (1957) distingue dos tipos de situaciones en las que se observa que los insectos producen sonidos. Por un lado, están aquellos “cantos solitarios” producidos por los machos adultos de manera más o menos continua y que son inhibidos por la presencia de otros organismos. Respecto de estos cantos, Alexander afirma que:
Funcionan al acercar a los machos y hembras adultas (al menos en algunas especies), ya sea por (1) hacer que la hembra se mueva hacia los machos (grillos), (2) hacer que la hembra haga un sonido que atraiga a la macho y le permite localizarla (saltamontes con cara inclinada), o (3) causando la congregación de un gran número de machos y hembras (cigarras). El conocimiento de estas funciones ha causado que nombres tales como “llamadas” y cantos “congregacionales” se apliquen a estos sonidos (1957, p. 102, mi énfasis).
Nótese aquí el uso explícito de la palabra “función”. La “función” que se atribuye al canto de “llamada” de los grillos es un efecto especial del mismo: hacer que la hembra se mueva hacia los machos. La “función” que se atribuye al canto “congregacional” (de las cigarras) es causar la agrupación de machos y hembras. Nótese además que es en virtud de poseer tales efectos que los cantos son clasificados: se los denomina “llamadas” o cantos “congregacionales” debido a tener ese efecto especial denominado “función”. En relación con los efectos sobre el comportamiento de la hembra que produce el canto de llamada, Alexander afirma que:
Este sonido afecta el comportamiento de una hembra sexualmente receptiva en los siguientes aspectos: 1) ella se orienta hacia la fuente del sonido con un alto grado de precisión y 2) ella se mueve de una manera constante en una línea bastante recta (si no hay obstrucciones) hacia la fuente del sonido (Alexander, 1960, p. 63).
Posteriormente, este comportamiento de orientación guiado por el sonido se denominará “fonotaxis”. En este sentido, podría decirse que la función del canto de llamada es producir la fonotaxis en las hembras sexualmente receptivas.
Por otro lado, están aquellos cantos que se producen como resultado de la presencia o actividad de otros organismos, y son clasificados en cuatro categorías:
(1) los producidos por machos, hembras o inmaduros como resultado de la presencia o actividades de animales de otras especies -molestarlas, capturarlas o sujetarlas- y que se denominan “gritos de alarma”, llamadas de “socorro” , sonidos de “protesta”, o sonidos “congregacionales”; (2) los producidos por un macho en presencia de una hembra de la misma especie, generalmente llamados cantos de “cortejo” o “apareamiento”; (3) los producidos por un macho en presencia de otro macho de la misma especie, generalmente llamado de “advertencia”, “intimidación” o “lucha”; Y (4) los producidos por una hembra en presencia de un macho de la misma especie (Alexander, 1957, p. 102).
Esta clasificación de los sonidos es resumida por Alexander por cuadros como el siguiente:
Figura 3: Secuencias comportamentales asociadas a los sonidos de los insectos (Elaboración propia, a partir de Alexander, 1957).
En el cuadro aparecen, de izquierda a derecha, el orden de insectos involucrado en un determinado comportamiento (por ejemplo, Orthoptera), la categoría de sonido en cuestión (por ejemplo, “llamada” [calling]), el tipo de organismo que responde a él (por ejemplo, hembra [female]) y con qué actividad (por ejemplo, aproximación [approach]) y aquello que “facilita” (por ejemplo, copulación [copulation]).
Nótese que, en el cuadro anterior, la “llamada” de los ortópteros también puede ser respondida por un macho [male], quién o bien se aleja [avoidance], o bien se aproxima [approach], dando lugar a una lucha [fight] que culmina en la separación de los machos [separation of males]. Todos estos efectos del canto, indica el cuadro, facilitan el “espaciamiento de las parejas reproductivas” [spacing of breeding pairs]. Entonces, el canto de llamada pareciera tener, según Alexander, dos funciones: por un lado, el canto de llamada posee la función de atraer a las hembras, facilitando la copulación, y, por otro lado, posee la función de separar a los machos, facilitando el espaciamiento de las parejas reproductivas. Todos estos son efectos del canto que, por alguna razón, son importantes para el autor. Alexander explicita estas funciones del canto de llamada en el siguiente pasaje:
Aunque la función básica del canto de llamada de los machos es en todo caso la atracción de hembras sexualmente receptivas, parece probable que en la mayoría de las especies estos sonidos tengan una mayor variedad de efectos sobre otros machos que los oyen que sobre las hembras. Así, parece que en diferentes situaciones el canto de un macho puede hacer que otro macho se vuelva más activo, se vuelva menos activo, se mueva hacia él, se aleje de él, empiece a cantar, pare de cantar, o cante a una tasa o ritmo diferente de lo que sería si no pudiera escuchar el primer canto masculino. En los Orthoptera y Cicadidae, la elaboración de estos diferentes efectos “colaterales” parecería en todos los casos depender del mejoramiento definitivo de la función primaria de aproximación de los sexos, de si esta mejora opera directamente o más o menos indirectamente a través de ajustes en la organización social de las especies (Alexander, 1960, p. 50, mi énfasis).
Así, según Alexander, la “función primaria” del canto de llamada es la atracción de hembras, lo cual facilita la copulación de los sexos. Los denominados “efectos colaterales” del canto, como el espaciamiento de los machos, pueden tener una función si mejoran aquella “función primaria” que es, fundamentalmente, reproductiva. La secuencia de comportamientos y efectos relevantes (en relación con “los ciclos reproductivos” de estos organismos) es representada por Alexander en esquemas como el de la figura 4 (infra).
Figura 4: Los ciclos reproductivos en Gryllus (Elaboración propia a partir de Alexander, 1962).
Sin embargo, esos no son los únicos efectos que tiene el canto de llamada. De hecho, Alexander (1967) nota cierta dificultad al intentar clasificar las funciones de los sonidos de los insectos en general, en tanto…
Cualquier señal puede tener un efecto sobre los miembros de otras especies, tanto sin significado selectivo o de un modo que tendrá (al menos inicialmente) un significado selectivo sólo con respecto a la especie responsiva (por ejemplo, los predadores pueden ser atraídos por las señales sexuales de las especies presas).
Para ilustrar algunas de esas complicaciones, existe evidencia de que bajo diferentes circunstancias los sonidos de llamada de los grillos machos pueden (a) atraer predadores; (b) atraer hembras; (c) actuar como un depresor de la locomoción (a altas intensidades) para las hembras; (d) causar chirridos agresivos y otras acciones agresivas; (e) estimular el llamado de otros machos; (f) inhibir la llamada de otros machos; y (g) causar que otros machos se alejen (Alexander, 1967, p. 499, mi énfasis).
Dado que el canto de llamada posee una gran cantidad de efectos conocidos, es interesante preguntarse por qué uno de ellos (la atracción de hembras conespecíficas) es aquel que es destacado como su función (primaria). Una respuesta posible, en este contexto, es que dicho efecto es importante en tanto es aquel que facilita la reproducción de estos organismos. En el próximo capítulo abordaré con mayor detalle la factibilidad de esta afirmación.
Por otra parte, Alexander (1957, 1960) nota que existen diferencias significativas entre los sonidos emitidos por un mismo individuo en diferentes situaciones naturales, por un lado, y los sonidos emitidos por individuos de diferentes especies, por otro. Todo ello sugiere, para Alexander, que podría existir una “capacidad discriminatoria” en los sistemas auditivos de las hembras que les permitiría hallar a los machos cantores de su propia especie.
El alto grado de diferenciación entre los sonidos producidos por un insecto determinado en diferentes situaciones, y su consistencia, sugiere que las diferencias estructurales en estos sonidos son conductualmente significativas. Del mismo modo, el hecho de que no se haya demostrado que dos especies de insectos, no importa cuán estrechamente relacionadas, tengan llamadas inseparables o cantos congregacionales, sugiere la probabilidad de capacidad discriminatoria (1957, p. 110).
Observaciones adicionales indican que las diferencias estructurales en los cantos de llamada de diferentes especies de insectos pueden actuar como mecanismos de aislamiento de especies. En el este de Estados Unidos, donde cientos de individuos de docenas de especies diferentes pueden estar cantando simultáneamente en un pequeño bosque o pantano, la incapacidad de las hembras para reconocer los cantos de los machos de su propia especie resultaría en una confusión total (…) Si esa hembra respondiera igualmente a todos los sonidos de los insectos de la zona, o incluso al sonido más intenso en su vecindad, su posibilidad de conocer a un macho de la misma especie se reduciría mucho y el tiempo requerido haría que esto fuera un sistema de comunicación ineficiente (1957, p. 111).
Si una señal particular es la principal responsable de la formación de parejas en cualquier grupo de especies, entonces las diferencias entre las señales de las especies que viven juntas evitarían gastar tiempo y energía en interacciones interespecíficas perjudiciales (1967, p. 509).
Independientemente de si el cruzamiento o hibridación de especies es perjudicial para los individuos, y el propio Alexander duda de ello (1967, p. 509), el hecho relevante es que las diferencias existentes entre los cantos del repertorio de un mismo individuo, y entre los cantos de llamada de diferentes especies, sugiere que las hembras tendrían la capacidad de distinguir entre tales cantos. Como veremos en el siguiente apartado, esta sugerencia fue importante para el estudio de las bases neurales de la fonotaxis. Nótese además que Alexander menciona que tales diferencias en los cantos serían indicativas de que los cantos de llamada podrían operar como un mecanismo de aislamiento reproductivo de las especies. En el siguiente capítulo (sección 5.1.1) trataré la influencia que podría haber tenido esta sugerencia dentro de la atribución de la función al canto de llamada.
4.4. La fonotaxis
Como vimos, la función del canto de llamada es la atracción de hembras conespecíficas (de la misma especie). Las hembras, ante la recepción del canto de llamada específica de su especie, ejecutan un comportamiento orientativo guiado por el sonido denominado “fonotaxis”. Que la función del comportamiento fonotáctico es el hallazgo de un macho conespecífico puede leerse en afirmaciones como la siguiente:
Para muchos animales la audición direccional es fundamental para su estilo de vida, ya que constituye la base de la detección de presas, la evitación de predadores o la localización de parejas (…) Los grillos en particular son un sistema neuroetológico modelo para el procesamiento auditivo y la orientación en los insectos, en tanto las hembras utilizan claves acústicas de los cantos de llamada de los machos para hallar parejas (Schöneich & Hedwig, 2010, p. 1, mi énfasis).
Utilizando un lenguaje comunicacional, se suele describir a este fenómeno como un sistema de comunicación en el cual los cantos de los machos son “señales” que deben ser “decodificadas” por el “receptor”, la hembra.
Para el hallazgo de un compañero intraespecífico y la reproducción, el emisor y el receptor tienen que ser emparejados [be matched] vía las señales acústicas. La información codificada en las señales acústicas del emisor debe ser decodificada por el receptor (Huber, 1990a, p. 318, mi énfasis).
Nótese aquí nuevamente el uso del lenguaje funcional. “Para” el hallazgo de un macho de su especie, la hembra (el receptor) “debe” decodificar la señal acústica del macho (el emisor). Dado que las hembras son atraídas por los cantos de los machos de su propia especie (cf. sección 4.4.1), se asume que dicha decodificación sería exitosa si la hembra logra resolver dos tareas:
El sistema nervioso del receptor debe cumplir dos tareas igualmente importantes: debe ser capaz de discriminar los cantos conespecíficos de otros sonidos bióticos y abióticos (reconocimiento del canto) y debe localizar la posición del emisor (localización del sonido) (Huber, 1990a, p. 318, mi énfasis).
De este modo, se asume que el procesamiento de la señal que da lugar a la fonotaxis implica que el sistema nervioso de las hembras debe ser capaz de resolver dos tareas: (1) el reconocimiento del canto específica de su especie y (2) la localización de su emisor. Si tales procesos son llevados adelante por el mismo o por distintos circuitos neurales y, en todo caso, cómo interactúan en el control de la fonotaxis, no es aún algo resuelto (Doherty, 1991). Sin embargo, a los fines de comprender los mecanismos neurales subyacentes a la fonotaxis, se han propuesto distintos modelos que pretenden dar cuenta de la manera en la que el sistema nervioso resolvería cada una de esas tareas.
En las siguientes secciones, abordaré el modo en que se indagaron las bases neurales de la fonotaxis en lo que respecta a la tarea de reconocimiento del canto conespecífico (1), dejando de lado, por razones de espacio, las indagaciones correspondientes a cómo resuelven la tarea de localización (2). De cualquier manera, será suficiente, para los fines del presente trabajo, profundizar sólo este aspecto de la investigación. Para ello, en la sección 4.4.1 exhibiré el tipo de estudios comportamentales llevados a cabo para indagar qué propiedades del canto de llamada son las relevantes para provocar la respuesta fonotáctica. Como veremos, dichos estudios fueron necesarios para investigar cuáles son las bases neurofisiológicas de dicha respuesta. Luego, en la sección 4.4.2 expondré uno de los modelos desarrollados para explicar cómo la hembra resuelve la tarea de reconocimiento del canto específica de su especie.
4.4.1. Los parámetros atractivos del canto de llamada
La primera demostración experimental de que las hembras de grillo pueden aproximarse a un macho distante utilizando sólo su canto como clave fue efectuada por Regen (1913), quien mostró que una hembra podía aproximarse a un receptor telefónico transmitiendo el canto de un macho que estridulaba en otra habitación. Inspirado directamente por las observaciones de Alexander, Walker (1957) mostró experimentalmente que dicho comportamiento es selectivo a los cantos de los machos de la misma especie. Walker realizó esta demostración estimulando a las hembras con cantos de machos de la misma especie y de machos de especies simpátricas[3], quienes respondieron mucho más fuertemente ante el primer estímulo. Más aún, Walker notaba que la tasa de pulsos (o sílabas) era la característica que difería mayormente entre las especies que compartían el mismo hábitat, y que tanto la tasa de pulsos como la preferencia de las hembras variaban juntamente con la temperatura del ambiente, sugiriendo que la tasa de pulsos podría ser la base de la especificidad de la respuesta. En este contexto, surge la siguiente pregunta: ¿qué características de los cantos son atractivas para las hembras? esto es, ¿qué parámetros del canto de llamada son relevantes para provocar la respuesta fonotáctica? Este tipo de pregunta apunta a especificar el estímulo señal o releaser que provoca el patrón de respuesta estudiado (cf. sección 2.3), y su abordaje involucró la realización de una serie de estudios experimentales que utilizaron variaciones artificiales de los patrones temporales naturales de los cantos (modelos o dummies, cf. sección 2.3), y midieron las respuestas de las hembras como síntoma de su preferencia.
Típicamente, en estos experimentos las hembras son colocadas en un laberinto-Y (por ejemplo, en Popov, Shuvalov, Svetlogorskaya, & Markovich, 1974; Popov & Shuvalov, 1977) o en un sistema de compensación de locomoción de Kramer (por ejemplo, en Doherty, 1985a, 1985b; Schmitz, Scharstein, & Wendler, 1983; Weber et al., 1981), cuyo nombre evoca a su inventor. El Laberinto-Y consiste en un carril bifurcado sobre el que se coloca al animal. En los extremos de cada brazo del laberinto se encuentran dos altavoces que reproducen alternativa (esquema sin elección) o simultáneamente (esquema con elección) distintos patrones de sonidos, para luego observar la preferencia de la hembra.
Por otra parte, el compensador de Kramer consiste en una esfera hueca sobre la cual el insecto camina libremente. Su movimiento es detectado mediante un sistema de escaneo por luz infrarroja que se conecta con un sistema de motores compensatorios, los cuales mueven la esfera en la dirección opuesta a la dirección en la que se mueve el insecto. Este movimiento compensatorio mantiene a la hembra en la cima de la esfera, permitiendo el estudio de su respuesta fonotáctica manteniendo el campo de sonido estable. De la misma manera que en el laberinto-Y, el insecto es expuesto a patrones acústicos con variaciones sistemáticas, en donde la preferencia de la hembra es medida como la probabilidad y/o el vigor con el que realiza la fonotaxis. De este modo, mediante el análisis de la velocidad y la dirección del movimiento, el rendimiento de la hembra en la tarea de rastreo puede ser cuantificado calculando el porcentaje de tiempo que el estímulo presentado es seguido (para más detalles acerca de este diseño experimental, véase (Weber, Thorson y Huber, 1981).
Los estímulos auditivos presentados a las hembras consisten en variantes artificiales de los cantos de llamada naturales que aíslan un posible parámetro atractivo. Los cantos de llamada naturales de los grillos de campo consisten en trenes de cuatro pulsos/sílabas, de una duración de 15-20mseg cada una, moduladas a una frecuencia portadora de 5kHz y repetidas a una frecuencia de 30Hz. Tales conjuntos de sílabas conforman un único chirrido, que se repite a una tasa de 2-3 chirridos por segundo. Las variaciones artificiales consisten, por ejemplo, en trenes de puras sílabas sin diferenciación de chirridos individuales, en tonos puros sin diferenciación de sílabas, en secuencias de sílabas con diferentes tasas de repetición o de duración, etc. (ver fig. 5, infra).

Figura 5: Propiedades temporales de cantos de llamada sintéticos. Parte superior: chirrido [chirp] de 4 sílabas, CD – duración del chirrido, CP – periodo del chirrido, SP – periodo de la sílaba, SD – duración de la sílaba. Parte intermedia: trino continuo [trill] sin diferenciación en chirridos individuales. Parte inferior: ráfaga de tono [tone burst] sin estructura silábica (Elaboración propia a partir de Doherty, 1985b).
De este modo, los experimentadores pudieron testear distintas hipótesis respecto de qué propiedades del canto natural son aquellas que provocan la respuesta fonotáctica. Así, por ejemplo, si la cantidad de sílabas por chirrido fuese un parámetro relevante de los cantos naturales, se espera que las hembras respondan con mayor probabilidad a estímulos dentro del rango de la cantidad de sílabas de los cantos naturales (3-5) que a estímulos fuera de ese rango.
De manera general, los resultados experimentales concuerdan en que el parámetro más importante de los cantos de llamada de los grillos de campo es la tasa de sílabas por segundo, la cual es de 30Hz, moduladas a una frecuencia portadora de 5kHz. La versión más extrema de esta hipótesis ha sido formulada por Thorson, Weber & Huber (1982), quienes sostuvieron la llamada “hipótesis de los 30Hz”. Según esta hipótesis, la modulación de las sílabas a una tasa de repetición a 30Hz es la única característica requerida para la provocación de la fonotaxis, siempre que la frecuencia de sonido sea audible para el insecto (entre 4 y 6 kHz). Si bien se ha comprobado (Doherty, 1985b) que dicho parámetro no es necesario ni suficiente para la fonotaxis, y que otros parámetros pueden ser relevantes, tales como la duración de las sílabas y del chirrido, la tasa de sílabas por segundo es considerada el parámetro del canto con mayor peso en la producción de la fonotaxis (Popov y Shuvalov, 1977; Weber, Thorson y Huber, 1981; Schildberger, 1984; Huber y Thorson, 1985; Huber, 1990a; Hedwig, 2006).
La importancia de estos estudios para la indagación de las bases neurales de la fonotaxis ha sido clave, por dos razones principales. Por un lado, tales estudios permitieron hallar los parámetros relevantes del canto de llamada para la producción de la fonotaxis, lo cual, veremos, contribuyó en la indagación experimental de las bases neurales de dicha responsividad. En palabras de los propios investigadores,
Tales estudios de comportamiento son necesarios para identificar las características del canto de llamada que influencian la fonotaxis de las hembras en orden de que uno sea capaz de correlacionar aquellas actividades neurales que reflejan los componentes comportamentalmente significativos del canto (Stout & Huber, 1981, p. 200).
Esto es, conocer los parámetros relevantes del canto permite correlacionar la actividad de ciertas neuronas con dichos parámetros, posibilitando el hallazgo de las bases neurales del procesamiento de la señal.
Por otro lado, tales estudios establecieron la base conductual sobre la cual contrastar distintas hipótesis acerca del modelo neural subyacente a la fonotaxis. En palabras de Horseman y Huber:
Una prueba útil de nuestra comprensión de las bases neurales de un comportamiento es producir modelos que combinen las características fisiológicas y la conectividad sináptica de las neuronas en la red subyacente con diversas suposiciones simplificadoras. La salida de un modelo se puede comparar con el comportamiento real para comprobar si las propiedades de red conocidas pueden dar cuenta de ello (Horseman & Huber, 1994, p. 399).
En definitiva, la importancia de estos estudios es doble, en tanto permitieron indagar experimentalmente las bases neurales de la fonotaxis, a la vez que permitieron contrastar los modelos neurales propuestos sobre la base de la selectividad exhibida por las hembras. En el próximo apartado veremos cómo este tipo de estudios contribuyeron a indagar las bases neurales del reconocimiento del canto conespecífico.
4.4.2. Las bases neurales de la fonotaxis: Un modelo para el reconocimiento del canto de llamada
En los grillos de campo, los órganos auditivos u “órganos timpánicos” se hallan localizados en sus patas delanteras, y los aferentes auditivos terminan en un ganglio denominado ganglio protorácico [prothoracic ganglion]. Las respuestas auditivas son llevadas del ganglio protorácico hacia el cerebro por medio de una única interneurona ascendiente, denominada AN1 (Schildberger, 1984) o TH1-AC1 (Kostarakos y Hedwig, 2012).
Dado que hasta este nivel de procesamiento no se hallaron neuronas selectivas a los parámetros de sonido relevantes para la fonotaxis (Wohlers y Huber, 1978, 1982; Schildberger, 1984; Huber, 1990a), es de esperarse que dicha selectividad se encuentre localizada a nivel cerebral.
Puedo haber dado la impresión de que tales células nerviosas ascendentes [AN1] en la vía auditiva del grillo son suficientes o incluso necesarias para la identificación de los sonidos conespecíficos, en tanto proveen la información requerida en los dominios de frecuencia, intensidad y tiempo. Pero un factor crucial debe ser tenido en mente. Ciertos parámetros críticos como la tasa de repetición de sílabas y la duración de los chirridos son señalizados por tales neuronas, pero ellas codifican de un modo igualmente fiel los diferentes arreglos temporales de otros cantos. Esto significa que la discriminación debe ocurrir a un nivel más alto, en el cerebro (Huber, 1978, p. 978).
Por lo tanto, un problema principal en el estudio de la vía auditiva del grillo es mostrar si el cerebro contiene neuronas identificables con propiedades de filtro temporal que de hecho coincidan con los parámetros de reconocimiento del canto conespecífico demostrados en el comportamiento (Schildberger, 1984, p. 172).
Para explicar el modo en el cual el cerebro de la hembra resuelve la tarea de reconocimiento del canto conespecífico, se propusieron diversos modelos orientados a delinear el posible mecanismo neural subyacente. Tales modelos intentan explicar el modo en el cual el sistema nervioso logra ser sensitivo a ciertos parámetros temporales de los cantos, esto es, aquellos que se mostraron que son relevantes en la producción de la fonotaxis. Con este objetivo, se apeló a diferentes conceptos. Algunos modelos apelan a un “modelo interno” de canto con el cual la señal recibida es comparada (por ejemplo, Clemmens & Hennig, 2013; Hennig, 2003; Hoy, 1978; Pollack & Hoy, 1979). Según esta idea, el patrón de pulsos es reconocido cuando sus características coinciden con las del modelo interno. Otros modelos apelan a un “filtro temporal” que sería resultado de la actividad de tres tipos de neuronas: las neuronas “filtro paso alto”, las neuronas “filtro paso bajo”, y las neuronas “filtro de paso de banda” o “de reconocimiento”, cuya actividad es determinada por la actividad de las primeras dos, formando un sistema similar al de las puertas AND (Schildberger, 1984, 1985; Huber, 1990a) El modelo que tomaré aquí es el modelo de “autocorrelación” de Hedwig, Kostarakos y Schoneich (Kostarakos y Hedwig, 2012, 2014; Schöneich, Kostarakos y Hedwig, 2015), en tanto es el modelo más actual y que cuenta, aparentemente, con mayor evidencia en lo que respecta a su implementación neural (Kostarakos y Hedwig, 2014).
Figura 6: modelo de “autocorrelación” propuesto para el reconocimiento del periodo de sílabas específico de la especie (Schoneich et al., 2015).
El modelo de “autocorrelación” apela, básicamente, a un procesamiento de la señal acústica por una doble vía paralela: una vía directa y una vía de retardo, las cuales convergerían en un detector de coincidencias (ver fig. 6). Según este modelo, el reconocimiento del patrón de pulsos se daría cuando el periodo de sílabas[4] de la señal recibida coincide con el retardo interno de la vía secundaria de procesamiento. De esta manera, si la respuesta a la primera sílaba de la vía de retardo coincide con la respuesta a la sílaba subsiguiente de la vía directa, el detector de coincidencias es activado, excitando, a su vez, un detector de características que responde selectivamente a la tasa de repetición de sílabas específica de la especie. A continuación, expondré este modelo con mayor precisión, y exhibiré la evidencia correspondiente a su posible implementación neural.
Con el fin de indagar el modo en el que el sistema nervioso de las hembras resuelve la tarea de reconocimiento del canto conespecífico, los investigadores analizaron la selectividad de las hembras a distintas características temporales de los cantos de llamada, y compararon dicha selectividad con la actividad de un conjunto de neuronas localizadas en el protocerebro anterior de estos insectos (detalles en Kostarakos & Hedwig, 2012).
El registro intracelular se enfocó en la vecindad de la región axonal de la interneurona AN1, en donde se expectaba que ocurran pasos cruciales del procesamiento auditivo. En dicha región se encontraron neuronas cerebrales locales (LN2, LN3, LN3*, LN4 y LN5) que respondían específicamente al canto de llamada. Estructuralmente, tales neuronas tienen en común el hecho de estar localizadas en el protocerebro anterior, con el cuerpo celular situado cerca del nervio óptico (ver figuras 7 y 8). Sus neuritas forman una estructura similar a un aro que se superpone con las arborizaciones axonales de AN1. Una de ellas (LN3*) tiene una proyección axonal hacia el lado contralateral[5] del cerebro, en donde repite el patrón de proyección en forma de aro. Contrariamente, las arborizaciones de LN2, LN3, LN4 y LN5 se encuentran restringidas al lado ipsilateral. A pesar de sus similitudes estructurales, tales neuronas muestran diferentes propiedades de respuesta.
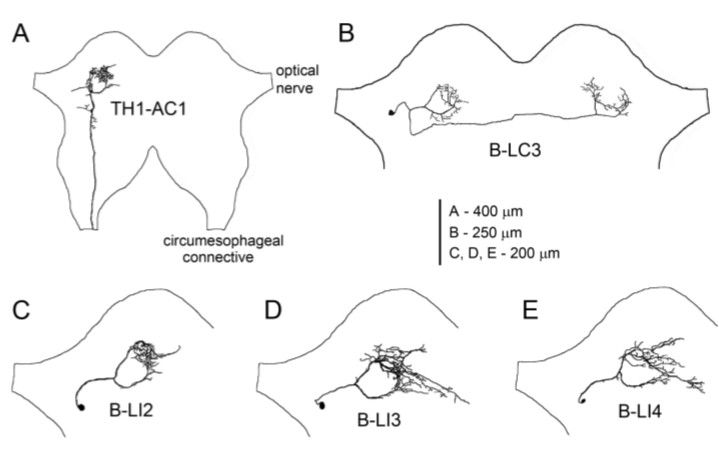
Figura 7: Estructura y localización de las neuronas auditivas cerebrales del grillo. A. Proyecciones axonales de la neurona ascendiente TH1-AC1 (AN1) en el protocerebro anterior. B. Morfología de B-LC3 (LN3*) con el cuerpo celular cercano al nervio óptico; sus arborizaciones dendríticas se superponen a los patrones de arborización en forma de aro ipsilaterales de TH1-AC1 (AN1) y las proyecciones axonales se superponen con los del lado contralateral. C-E Morfología de B-LI2 (LN2), B-LI3 (LN3) y B-LI4 (LN4) con el cuerpo celular lateral (Kostarakos y Hedwig, 2012).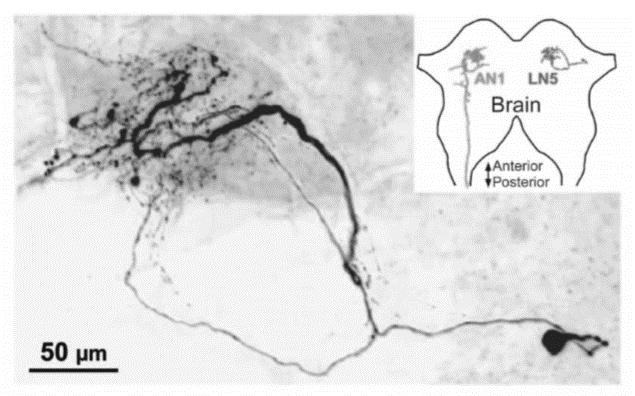
Figura 8: Estructura y localización de la interneurona cerebral LN5. Las arborizaciones dendríticas se encuentran en vecindad inmediata con las terminales de AN1 en el cerebro. En el recuadro superior sólo se representan las neuronas AN1 izquierda y LN5 derecha con el fin de indicar la localización de las neuronas en el cerebro (Schöneich, Kostarakos y Hedwig, 2015).
Dado que el comportamiento fonotáctico máximo ocurrió con determinados patrones temporales (una duración de pulsos de 15mseg, con un intervalo de pulsos de 15mseg y un periodo de pulsos de 34-42mseg) los investigadores indagaron la actividad de disparo [spike activity] de tales neuronas cerebrales ante diferentes patrones temporales, esperando hallar la fuente de dicha preferencia. La actividad exhibida por estas neuronas fue evaluada en relación con su respuesta ante tres modelos acústicos: el modelo de duración de pulsos (DP), el modelo de intervalo de pulsos (IP) y el modelo de periodo de pulsos (PP). En cada uno de ellos se presentaron estímulos con diferentes valores de DP, IP y PP, y se midieron las respuestas de las neuronas estudiadas mediante técnicas de registro intracelular. Los resultados pueden resumirse como sigue.
Ante todos los modelos temporales presentados, LN2 respondió con un potencial post sináptico excitatorio (PPSE) con una corta latencia, siguiendo el patrón de actividad de AN1. La actividad de ambas neuronas decrece gradualmente del primero al cuarto pulso de un chirrido, y sus actividades de disparo no mostraron selectividad ante ningún patrón de sonido. Dada la similitud en las respuestas de ambas neuronas, los investigadores infieren que es probable que AN1 envíe inputs excitatorios directos a LN2 (Kostarakos & Hedwig, 2012, 2014).
A diferencia de LN2, las neuronas cerebrales LN3, LN3* y LN4 mostraron ser más sensitivas a las características temporales de los cantos.
LN3 y LN3* mostraron poseer propiedades de respuesta similares, y por ello me referiré a ellas simplemente como LN3, tal como hacen los autores. Ambas respondieron máximamente a DP de 20mseg, IP de 15mseg y PP de 34mseg, coincidiendo altamente con la preferencia exhibida por el comportamiento fonotáctico de las hembras. Sin embargo, las respuestas exhibidas ante valores no óptimos no fueron del todo coincidentes con dicho comportamiento.
Llamativamente, cuando LN3 fue testeada con intervalos de pulsos (IP) menores a 10mseg o mayores a 35mseg (fuera del rango natural de los cantos de llamada), respondió ante cada pulso de sonido con dos respuestas similares y separadas, exhibiendo una despolarización gradual supra-umbral inicial (con disparos de potenciales de acción) y luego una despolarización sub-umbral retardada (sin disparos). Sin embargo, cuando fue testeada con pares de pulsos con intervalos de silencio de 15-25mseg, correspondiente al rango natural de intervalos de pulsos, LN3 mostró una respuesta diferente. En efecto, LN3 mostró una respuesta más fuerte ante el segundo pulso de sonido, en tanto la despolarización retardada al primer pulso de sonido coincidió con la despolarización inicial provocada por el segundo pulso. De acuerdo con los investigadores, estos datos sugieren que LN3 podría ser la primera neurona dentro de la red de procesamiento que muestra ser selectiva a los parámetros temporales del canto de llamada, en tanto responde con mayor intensidad a aquellos pares de pulsos que poseen intervalos de silencio dentro del rango de los cantos de llamada naturales. Más aún, los investigadores comprenden que LN3 cualifica como un “detector de coincidencias”, en tanto su selectividad está basada en la suma de inputs excitatorios directos y retardados (Schöneich et al., 2015; Kostarakos & Hedwig, 2014).
A diferencia de las neuronas hasta aquí presentadas, LN4 respondió a todos los modelos temporales con dos tipos de respuestas: una inhibición inicial (PPSI) seguida de una excitación (PPSE). Sus respuestas máximas se dieron a una DP de 15mseg, una IP de 20mseg y una PP de 34mseg, exhibiendo, como LN3, una fuerte coincidencia con la selectividad de la respuesta fonotáctica. Su respuesta a valores no óptimos dio lugar a PPSE que raramente superaron el umbral de disparo, y su actividad total fue altamente coincidente con la preferencia fonotáctica de las hembras.
El patrón de respuesta de LN4 ante los distintos valores de intervalos de pulsos (IP) es altamente relevante para el modelo de autocorrelación. En efecto, cuando los pulsos fueron presentados con intervalos de silencio menores a 10mseg o mayores a 50mseg (fuera del rango de IP natural), la excitación producida permaneció por debajo del umbral de disparo. Sin embargo, la presentación de un segundo pulso de sonido luego de 20mseg de silencio (un valor dentro del rango natural de los cantos de llamada) causó una despolarización supra-umbral que llevó a la generación de potenciales de acción. Globalmente, el análisis cuantitativo de la actividad de LN4 mostró ser selectiva a intervalos de pulsos de 15 a 25mseg. De acuerdo con los investigadores, estos datos muestran que LN4 es altamente selectiva a los patrones temporales del sonido, en tanto responde con inhibición a los patrones de pulsos con tasas de repetición bajas o altas, mientras que responde con potenciales de acción sólo dentro de un rango estrecho de intervalos de pulsos, esto es, a aquellos correspondientes al rango temporal de los cantos de llamada. Dadas estas características, los investigadores consideran a LN4 como un “detector de características”, que dispara únicamente cuando los pulsos son presentados con el intervalo correspondiente a los cantos de llamada específicas de la especie (Kostarakos & Hedwig, 2014; Schöneich et al., 2015).
Hasta aquí sabemos que, cuando el estímulo presentado tiene el intervalo de pulsos apropiado (15-20mseg), el “detector de coincidencias” (LN3) responde a ambos pulsos, pero con una respuesta más potente ante el segundo pulso de sonido. Como vimos, dicha respuesta depende de la suma de la respuesta directa al segundo pulso y la respuesta retardada al primer pulso. Según los investigadores, esto implica que el “detector de coincidencias” requiere un input retardado luego de alrededor de 40mseg, correspondiente al periodo de pulsos específico de la especie. Esto es, requiere un input retardado luego del periodo de tiempo correspondiente al intervalo de pulsos (~20mseg) más la duración del segundo pulso (~20mseg). Crucialmente, los investigadores hallaron que la actividad de la neurona LN5 podría ofrecer dicho input retardado a LN3.
LN5 fue catalogada como una “interneurona no disparadora” [nonspiking interneuron], en tanto no se han registrado potenciales de acción en ella, ni pudieron ser provocados mediante inyecciones de corriente en la membrana celular. Sin embargo, ante pulsos de sonido individuales, LN5 respondió generando una inhibición inicial seguida de una despolarización sub-umbral mediadas por un mecanismo conocido como Rebote post Inhibitorio (PIR). Específicamente, LN5 responde con una inhibición inicial que alcanza una hiperpolarización máxima a los 35mseg. Subsecuentemente, la membrana de LN5 se despolariza, alcanzando su despolarización máxima a los 43mseg. Asimismo, luego de una hiperpolarización de 20mseg inducida por una inyección de pulsos de corriente hiperpolarizante, LN5 generó igualmente una despolarización retardada. De este modo, durante la estimulación acústica, LN5 genera una respuesta excitatoria con un retardo correspondiente al periodo de pulsos específico de la especie (~40mseg). Así, el Rebote post Inhibitorio (PIR) de esta neurona coincide cercanamente con la excitación retardada del detector de coincidencias LN3. Para los investigadores, esto indica que LN5 envía la excitación retardada a LN3 y, por lo tanto, refieren a LN5 como la neurona de la “vía de retardo” de la red (Schöneich et al., 2015) (véase figura 9, infra, para un resumen de la actividad de las neuronas aquí tratadas).
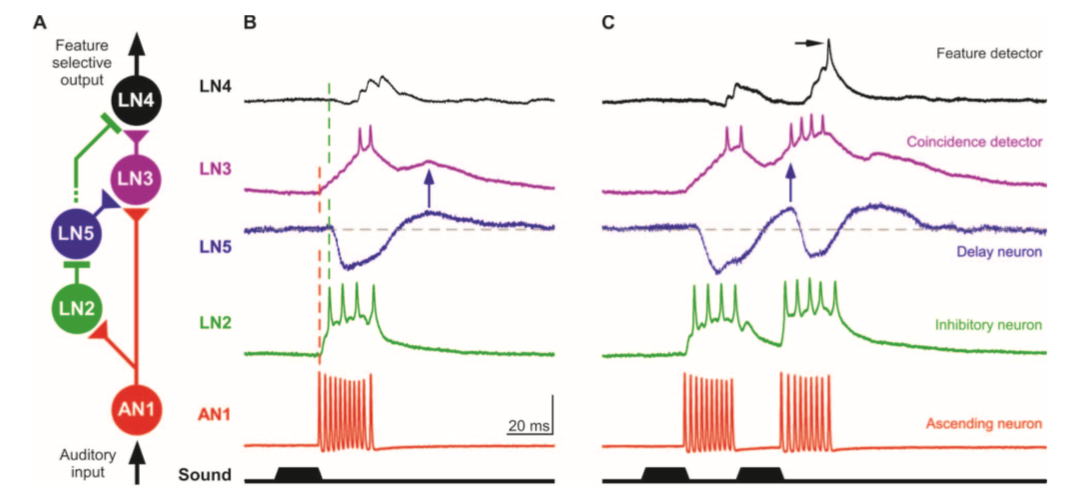
Figura 9: Implementación neural del modelo de “autocorrelación”. (A) Circuito neural basado en las propiedades de respuesta y la latencia de las respuestas neurales. Los triángulos y rectángulos indican sinapsis excitatorias e inhibitorias, respectivamente. (B y C) Las respuestas típicas de las cinco neuronas auditivas están alineadas al final de un único (B) o un par (C) de pulsos de sonido con 20mseg de intervalo. (B) El disparo de AN1 está seguido inmediatamente por una despolarización rápida de LN2 y una despolarización gradual de LN3. La inhibición en LN4 y LN5 sigue la actividad de disparo de LN2 con una corta latencia. El momento de la despolarización PIR de LN5 se corresponde con el EPSP retardado en LN3 (flecha azul). La actividad de disparo de LN3 precede a la excitación de LN4. (C) La respuesta de LN3 es aumentada por un segundo pulso de sonido presentado luego de un intervalo de 20mseg, al coincidir los inputs excitatorios directos (vía AN1) y retardados (vía LN5). Conducida por una fuerte actividad de LN3, LN4 ahora supera su inhibición y dispara (flecha negra) (Schoneich et al., 2015).
De este modo, el circuito neural subyacente al reconocimiento del canto conespecífico según el modelo de “autocorrelación” estaría conformado como sigue.
AN1 es la única interneurona ascendiente que lleva inputs auditivos al cerebro en respuesta al canto de llamada. Las latencias de respuesta y la frecuencia de sintonización de las neuronas cerebrales locales LN2 y LN3 indican, según los investigadores, que AN1 excita directamente ambas neuronas mediante PPSEs. Por ello, los investigadores cualifican a AN1 como la “vía directa” de la red de procesamiento, en tanto lleva las respuestas directas ante los pulsos de sonido a LN2 y LN3. Por su parte, LN2 parece conducir inputs inhibitorios a LN4 y LN5, que siguen su patrón de actividad. En la neurona LN5, dicha inhibición desencadena PIR, que lleva una despolarización retardada a LN3. De este modo, LN3 recibe dos inputs excitatorios: uno directo, enviado por AN1 (la “vía directa”), y uno retardado, enviado por LN5 (la “vía de retardo”). Cuando el intervalo de pulsos presentado es el específico de la especie, ambos inputs excitatorios coinciden, y LN3 dispara con mayor fuerza ante el segundo pulso de sonido. Por ello, los investigadores tratan a esta neurona como el “detector de coincidencias” de la red. La respuesta realzada del “detector de coincidencias” LN3 al segundo pulso de sonido proveería una fuerte excitación a LN4 (el “detector de características”) que supera la inhibición de LN2 y dispara.
En resumen, las propiedades de respuesta de las neuronas cerebrales y el ritmo secuencial de actividad que manifiestan apoyan una arquitectura neural en la cual AN1 provee una “vía directa” del patrón de pulsos, y LN2 y LN5 establecen una “vía de retardo”. Cuando la respuesta al segundo pulso de la “vía directa” coincide con la respuesta al primer pulso de la “vía de retardo” en el “detector de coincidencias” LN3, éste excita suficientemente al “detector de características” LN4, que así supera la inhibición de LN2 y dispara. Según el modelo de autocorrelación, esta es la manera en la que el sistema nervioso central de las hembras logra reconocer el canto de llamada correspondiente a un macho de su especie (en particular, el patrón de pulsos específico de su especie).
En este punto, considero que es posible apreciar el modo en el cual el lenguaje funcional también está involucrado en la indagación de las bases neurales de la fonotaxis. Creo que es razonable asociar el lenguaje funcional al modelo de “autocorrelación” presentado aquí. Esto es, creo que el lenguaje funcional utilizado en esta indagación se aplicó cuando se intentó generar un modelo que explique cómo la hembra resuelve la tarea de reconocimiento del canto conespecífico. Dicha tarea, se propone, se resuelve mediante el conjunto roles o funciones que constituyen al modelo de autocorrelación. Así, por ejemplo, la “vía de retardo” establece una función, esto es, un efecto que consiste en retrasar la excitación producida por cierto estímulo (el pulso de sonido) el tiempo correspondiente al periodo de sílabas específico de la especie. Este efecto es relevante en orden de producir, junto con las otras funciones, la salida del modelo de autocorrelación: el reconocimiento del canto de llamada conespecífico. La función “vía de retardo”, si el modelo ha de ser correcto, debe estar implementada de alguna manera por el sistema nervioso de la hembra. Dicha implementación se cree que fue hallada en las neuronas LN2 y LN5 y ciertas actividades que tales neuronas exhiben. LN2 inhibe LN5, y dicha inhibición desencadena PIR en LN5. De este modo, LN2 (y su efecto inhibitorio) y LN5 (y su efecto PIR), implementan la función “vía de retardo” del modelo.
Nótese que tales neuronas cualifican como la implementación de cierta función (la “vía de retardo”) en tanto exhiben cierta estructura y cierta actividad. La estructura (por ejemplo, la arborización de sus neuritas) y actividad (por ejemplo, PIR) de tales neuronas cobra relevancia en orden de explicar cómo es implementada la función “vía de retardo”. En este sentido, podría decirse que, por ejemplo, realizar “PIR” es la función de LN5 en tanto es dicha actividad, junto con la de LN2, la que explica cómo se implementa la “vía de retardo”.
Continuaré esta discusión en el próximo capítulo, en el cual abordaré con mayor detalle el uso del lenguaje funcional de la neuroetología. Para finalizar, realizaré un recuento de las cuestiones vistas en este capítulo que deberán ser retomadas en el siguiente.
4.5. Resumen
A lo largo de este capítulo me enfoqué en presentar la investigación neuroetológica de la comunicación acústica del grillo de campo, destacando el uso del lenguaje funcional involucrado en ella. Dicho lenguaje fue manifiesto en dos instancias específicas. En primer lugar, hemos visto que el lenguaje funcional se aplicó a ciertos comportamientos producidos por los insectos. Así, la función del “canto de llamada” de los grillos de campo fue especificada como la atracción de hembras, lo cual facilita la copulación (sección 4.2). Correlativamente, las hembras, ante los cantos de llamada de los machos ejecutan un comportamiento denominado “fonotaxis”, el cual tendría la función de hallar a los machos conespecíficos. Dado que los insectos producen distintos cantos según el contexto y según la especie, los investigadores infirieron que debe existir cierta “capacidad discriminatoria” en las hembras, las cuales serían atraídas sólo por los cantos conespecíficos. En segundo lugar, vimos que el lenguaje funcional fue aplicado cuando se abordó la cuestión sobre las bases neurales de la fonotaxis, esto es, las bases neurales de la capacidad de las hembras de guiarse por el canto de llamada de un macho conespecífico. Así, por un lado, se dijo que, para lograr hallar un macho de su especie, las hembras (o sus sistemas nerviosos) deben resolver dos tareas: (1) el reconocimiento del canto específica de su especie y (2) la localización de su emisor (sección 4.3). Como vimos, la utilización de modelos de cantos de llamada naturales y artificiales en distintos experimentos conductuales fue crucial para hallar aquellos parámetros del canto que son relevantes para la producción de la fonotaxis (en términos etológicos, el estímulo señal) y así avanzar sobre sus bases neurales (en etología, su mecanismo desencadenador) (sección 4.3.1). En este punto, el lenguaje funcional se utilizó cuando se generó un modelo (el modelo de “autocorrelación”) para explicar cómo los sistemas nerviosos de las hembras lograrían resolver la tarea de reconocimiento del canto en base a los parámetros hallados relevantes (sección 4.3.2). En este punto, los investigadores utilizaron el lenguaje funcional para describir los distintos componentes del modelo de autocorrelación, así como su posible implementación neural. De este modo, el modelo de autocorrelación estaría compuesto por distintas funciones: la “vía directa”, la “vía de retardo”, el “detector de coincidencias”, etc. Dichas funciones se hallaron implementadas por ciertas neuronas, en virtud de su estructura y ciertos patrones de actividad. De este modo, se explicaría cómo los sistemas nerviosos de las hembras son capaces de reconocer los cantos de los machos de su especie.
En el próximo capítulo intentaré clarificar el uso del lenguaje funcional explicitado en este capítulo apelando a los enfoques vistos en el capítulo 3.
- Por ejemplo Schwabe (1906), Sihler (1924) y Snodgrass (1926).↵
- El Dr. Richard Alexander (1930-actualidad) es profesor y curador de insectos emérito del Museo de Zoología de la Universidad de Michigan, EEUU. Su trabajo se abocó en buena medida al estudio de la biología y la sistemática de los grillos, saltamontes y cigarras. Recibió múltiples premios y reconocimientos tales como el Newcomb Cleveland Prize de la American Association for the Advancement of Science (AAAS) y la medalla Daniel Giraud Elliot de la National Academy of Sciences de Estados Unidos.↵
- Se dice que dos especies o poblaciones son simpátricas cuando poseen una distribución geográfica superpuesta, de manera que ocupan en mismo área y pueden encontrarse entre sí (Futuyma, 2013).↵
- El periodo de las sílabas se corresponde con el tiempo transcurrido entre el comienzo de una sílaba y el comienzo de la siguiente, incluyendo el intervalo de silencio transcurrido entre ambas. Este parámetro es recíproco con la tasa de sílabas por segundo, en tanto la cantidad de sílabas por segundo depende de la duración de las sílabas y sus intervalos. El grillo de campo posee un periodo de sílabas de 34-42mseg, con intervalos de silencio de alrededor de 20mseg, correspondiente a una tasa de repetición de sílabas de 30Hz.↵
- Se dice que dos ítems o estructuras se encuentran ubicadas del lado “ipsilateral” cuando están ubicadas del mismo lado con respecto a una línea media imaginaria. De un modo semejante, se dice que dos ítems o estructuras (en este caso, el cuerpo celular de LN3* y su proyección axonal) son “contralaterales” cuando se encuentran ubicadas en los lados opuestos de una línea media imaginaria.↵

