





Reflexiones en torno al caso de estudio
El objetivo de este capítulo es abordar la cuestión acerca del lenguaje funcional de la neuroetología. Aquí trataré la cuestión de cómo el concepto de función, y el lenguaje funcional en general, son utilizados por los neuroetólogos, qué significado parecen atribuirle y qué rol cumple en sus explicaciones acerca del comportamiento animal.
Dado que en este trabajo de tesis me enfoqué en un caso particular de la neuroetología, la estructura y el contenido del presente capítulo se encontrarán en estrecha relación con el capítulo anterior. Sin embargo, en este capítulo intentaré ofrecer reflexiones metateóricas referidas a dicho caso que permitirán ofrecer una evaluación de la adecuación de los distintos enfoques filosóficos presentados en el capítulo 3.
Siguiendo la estructura del capítulo precedente, entonces, en primer lugar examinaré la cuestión del lenguaje funcional aplicado al canto de llamada del grillo de campo (sección 5.1); en segundo lugar, trataré la cuestión del lenguaje funcional aplicado a las bases neurales de la fonotaxis en lo que respecta a la tarea de reconocimiento del canto conespecífico (sección 5.2); finalmente, ofreceré algunas conclusiones parciales referidas a este caso de estudio en particular (sección 5.3) y daré lugar a las conclusiones generales que serán presentadas en el último capítulo.
5.1. El lenguaje funcional en el estudio del comportamiento
Como vimos (sección 4.3), la “función primaria” que se atribuye al canto de llamada de los grillos de campo es un efecto particular de las mismas: la atracción de hembras conespecíficas, lo cual facilita la copulación de las parejas sexuales. Sin embargo, Alexander (1960) notaba que tales cantos también tienen efectos sobre otros machos, los cuales podían o bien alejarse, o bien acercarse y dar lugar a una lucha entre machos, todo lo cual finalizaba en la separación de las parejas reproductivas. Tales efectos, para Alexander, son funcionales si mejoran “la función primaria de aproximación de los sexos” (Alexander, 1960, p. 50). De este modo, desde Alexander, la función primaria de los cantos de llamada es la atracción de hembras conespecíficas, y una función, que podemos denominar secundaria, es el espaciamiento de los machos conespecíficos. Sin embargo, como vimos, tales funciones no son los únicos efectos que posee el canto de llamada. El canto de llamada puede tener también el efecto de atraer predadores, de causar chirridos agresivos en otros machos, de estimular el llamado de otros machos, etc. (Alexander, 1967). De este modo, una cuestión fundamental en la discusión acerca del estatus de las explicaciones funcionales y de la naturaleza del lenguaje funcional reside en explicar por qué ciertos efectos son privilegiados por sobre otros, atribuyéndoles el carácter de función.
Teniendo en cuenta los enfoques filosóficos presentados en el capítulo 3, intentaré evaluar su adecuación para la siguiente atribución funcional:
Canto de llamada:
- Función primaria: efecto de atraer hembras conespecíficas.
- Función secundaria: efecto de espaciar los machos conespecíficos.
Con el fin de aplicar y evaluar la adecuación de tales elucidaciones filosóficas a este caso particular, en el siguiente apartado trataré la relación entre el lenguaje funcional y la evolución por selección natural, abordando así la adecuación del enfoque etiológico y de la versión etiológica del enfoque de selección no histórica a este caso de estudio. Luego, en el apartado 5.1.2. trataré la manera en la que se determinó la atribución funcional en cuestión, abordando la adecuación del enfoque sistémico, de valor de supervivencia y de selección no histórica (en su versión de valor de supervivencia).
5.1.1. Función y evolución
Con el fin de aplicar y evaluar la adecuación del enfoque etiológico, recordemos en primer lugar qué características posee la atribución funcional para este enfoque. Como vimos (sección 2.4) la atribución de una función “propia” depende de la posesión de evidencia evolutiva que permita establecer la acción de la selección natural sobre el ítem funcional en organismos ancestrales. Esto es, la atribución funcional depende de la identificación de aquel efecto pasado del ítem que fue seleccionado (Millikan, 1984, 1989) o que contribuyó al fitness inclusivo de los organismos ancestrales que lo poseían (Neander, 1991a, 1991b), y que explica la presencia del ítem en los organismos actuales. Nótese que estas afirmaciones también son consistentes con el enfoque de selección no histórica (sección 2.5) en su versión etiológica, esto es, cuando se aplica a un régimen selectivo pasado (Walsh, 1996).
De este modo, la atribución funcional del canto de llamada, según la perspectiva etiológica, sería equivalente a la siguiente afirmación:
Enfoque etiológico:
Función propia del canto de llamada: efecto pasado del canto de los organismos ancestrales que fue seleccionado/que contribuyó al fitness inclusivo de tales organismos y que explica por qué los cantos de llamada se encuentran presentes en los organismos actuales.
Dado que, como vimos, el canto de llamada posee al menos dos funciones (la atracción de hembras y el espaciamiento de machos), es difícil establecer cuál de ellas, según el enfoque etiológico, sería aquella que sería la “función propia” del canto de llamada. Para establecer ello habría que incurrir en el estudio evolutivo que, precisamente, mostraría cuál de tales efectos pasados es aquel que ha sido seleccionado y que explica la presencia actual del rasgo.
Si bien es una cuestión compleja establecer con precisión qué tipo de evidencia poseía Alexander para atribuir las funciones al canto de llamada, en tanto sus estudios involucran una gran cantidad de datos heterogéneos, tales como evidencia observacional, experimental y comparada, en la próxima sección veremos que la evidencia evolutiva requerida por el enfoque etiológico no fue necesaria para determinar la función del canto de llamada. Esto es, la atribución funcional no fue dependiente de la identificación de aquel efecto pasado que contribuyó al fitness inclusivo de los organismos ancestrales, y que explica la presencia actual del canto de llamada en los organismos actuales. De hecho, como mostraré a continuación, los estudios acerca de la función del canto de llamada no poseían como objetivo el establecimiento del origen evolutivo del rasgo, sino que, por el contrario, dichos estudios funcionales constituyeron parte de la evidencia que se utilizó para determinar el origen evolutivo. Esto puede apreciarse al examinar qué tipo de evidencia fue relevante a la hora de indagar la historia evolutiva del canto de llamada y la posible acción selectiva pasada sobre ella.
En primer término, es preciso destacar que los cambios evolutivos y la acción selectiva sobre el comportamiento suelen considerarse difícil de establecer, en tanto, se dice, la evidencia paleontológica de especies ancestrales no se encuentra disponible para rasgos comportamentales, fundamentalmente porque “las acciones no se fosilizan” (Otte, 1992, p. 25). A falta de este tipo de evidencia, las generalizaciones acerca de la evolución del comportamiento suelen estar basadas en comparaciones sistemáticas entre taxones actuales. En relación a nuestro caso particular,
Obviamente no existen fósiles de los cantos de los grillos, y toda la evidencia debe provenir de estudios comparados de especies modernas, suplementada por lo que sabemos acerca de las relaciones entre los grillos desde los fósiles ortópteros (Alexander, 1962, p. 443).
La evidencia de que la evolución de la comunicación de los grillos ha tomado cursos particulares proviene de una comparación de más de 700 especies grabadas en cintas durante los últimos 20 años principalmente por Walker, Alexander, y Otte en Norteamérica, Australia, África y varias islas del Pacífico y el Caribe (Otte, 1992, p. 25).
Basado en este tipo de evidencia, Alexander (1962) afirma que todos los grillos estridulantes (alrededor de 2000 especies) deben ser descendientes de una misma especie que vivió hace alrededor de 150 millones de años. Las razones que otorga para creer ello pueden resumirse como sigue:
- Todos los fósiles y los grillos modernos poseen el mismo aparato estridulatorio y el mismo aparato auditivo, y tales aparatos son compartidos con los Tettigoniidae (saltamontes longicornios), una familia que, según indicios fósiles, se habría separado de la línea de los grillos durante el periodo Jurásico.
- Todos los grillos estridulantes modernos producen los sonidos intensos que sólo los grillos son capaces de producir debido a su dispositivo estridulador.
- La función de llamada está muy extendida y es muy similar entre los Gryllidae y Tettigoniidae, indicando que habría aparecido antes de que las dos familias se conviertan en líneas evolutivas separadas.
Nótese que (1) y (2) es evidencia de datos comparados estructurales, en tanto compara los aparatos estridulatorios y los sonidos de diferentes organismos actuales y pasados. (3) es evidencia comparada funcional, en tanto compara las funciones que cumplen los cantos de llamada en distintos tipos de organismos. Es decir, la atribución funcional se encuentra presupuesta en el estudio evolutivo, y no viceversa.
En virtud de las razones brindadas, para Alexander, la primera señal acústica en los grillos no habría sido una señal “de los grillos”, en tanto apareció en un ancestro común de los Gryllidae y los Tetiigoniidae. Dicha primera señal, para Alexander, habría tenido lugar como mediadora del cortejo, por las siguientes razones:
- El primer sonido que realmente tuvo importancia auditiva habría tenido baja intensidad y habría sido operativo sólo entre individuos en cercana proximidad, debido a que los aparatos estridulatorios y auditivos elaborados de los tetigonideos modernos no podrían haber aparecido completamente [full-blown].
- El levantamiento de las alas y el aleteo como estímulo visual y auditivo, y también en relación con la exposición de las glándulas dorsales, ocurre no sólo entre muchas subfamilias de grillos, sino también entre los tetigonideos (saltamontes) y los blatodeos (cucarachas) (que habrían divergido de otros ortópteros durante el Paleozoico). Esto es, la vibración (silenciosa o sonora) de las alas delanteras ocurre actualmente durante el cortejo en los machos de todos los ortópteros en los cuales las hembras montan activamente al macho en la copulación.
Nótese el carácter de esta evidencia. (1) es un dato del (posible) estado ancestral del rasgo, esto es, que habría sido de baja intensidad y operativo a distancias cercanas. (2) es evidencia comportamental comparada, en tanto surge de la comparación del comportamiento de cortejo en distintas familias de insectos emparentados. Este dato es importante en tanto establece que el movimiento de las alas (la base de la estridulación), tanto sonoro como silencioso, se encuentra presente en el comportamiento de cortejo de organismos actuales emparentados filogenéticamente y que, por lo tanto, habría estado presente en el cortejo del ancestro común de dichos organismos.
En virtud de estos datos, Alexander afirma que la primera señal acústica (en la historia evolutiva de los sonidos de estos insectos) habría sido una señal de cortejo:
Podemos decir con cierta seguridad, entonces, que la primera señal en el sistema acústico del grillo (o de los tetigonideos) fue una señal de cortejo, y que durante el tiempo que los grillos se convirtieron en una línea evolutiva separada el sistema habría progresado o bien hacia un estado de dos señales o bien la señal original de cortejo habría sido transformada en una señal de llamada (Alexander, 1962, p. 451).
Con respecto a la evolución de la señal de cortejo hacia la señal de llamada, Alexander dice basarse en la siguiente evidencia:
- Con respecto a las interacciones macho-hembra, estas dos cantos difieren sólo cuantitativamente en su principal función: el canto de llamada atrae a la hembra desde distancias lejanas; el canto de cortejo la atrae desde una corta distancia.
- La diferencia funcional cuantitativa entre el cortejo y la llamada está relacionada con diferencias cuantitativas estructurales. En las especies con los cantos de cortejo más simples, los cantos de cortejo difieren de las de llamada principalmente en intensidad y duración. El macho trina continuamente cuando llama, y trina brevemente, con interrupciones erráticas, cuando corteja. La intensidad y la duración son características que afectan primariamente el rango y la redundancia, los dos atributos que tendrían que ser mejorados si una señal de corto alcance fuera transformada o ampliada para incluir una función exitosa de largo alcance.
- La llamada y el cortejo están relacionados fisiológicamente en el macho: un macho llamando está “preparado” [primed] para el cortejo (requiere menos estímulos precisos o elaborados para empezar a cortejar que en otras situaciones). Él no empieza a llamar hasta que está preparado para cortejar y copular y un macho cortejando pasará a llamar muy frecuentemente si su cortejo es interrumpido por la partida de la hembra.
Nótese nuevamente el carácter de esta evidencia. (1) establece una diferencia funcional (actual) entre los cantos de cortejo y los cantos de llamada, principalmente, la distancia o el alcance. (2) establece que dichas diferencias funcionales (actuales) se corresponden con ciertas diferencias estructurales (actuales) de los sonidos, esto es, la intensidad y la duración. (3) establece una relación (actual) entre el comportamiento de llamada y de cortejo, en tanto se influencian causalmente entre sí.
En virtud de dicha evidencia, Alexander afirma que:
Creo que hubo una selección por un incremento en la ritmicidad, intensidad y duración en el canto de cortejo original (una o muchas veces, esto hace poca diferencia) porque esas características mejoraban la función de cortejo en sí misma, mediante el incremento en la consistencia, rango y redundancia. Eventualmente, mediante este tipo de cambio, este canto debió haberse vuelto operativo a tales distancias que a veces era ventajoso (esto es, en algunas instancias o en algunas especies) para el macho estridular sin contacto con la hembra, y a veces era ventajoso para la hembra ser atraída al escuchar esos sonidos cuando no estaba en contacto con el macho de otro modo. De este modo, la función de llamada, en la forma aproximada que asume hoy en día, podría haber evolucionado. Pero adicionalmente, con el cambio en el desencadenante del canto del macho, el camino habría sido abierto para la evolución de dos diferentes señales que podrían seguir sus propios caminos separados. Podría sólo ser necesario para el macho desarrollar señales estructuralmente diferentes, con efectos ligeramente diferentes, para las dos situaciones diferentes; o (quizás originalmente) [fue necesario] para la hembra comenzar a responder diferentemente a la señal original que sirvió para el llamado y el cortejo, dependiendo de si o no ella estaba en contacto con el macho a través de otros sentidos que el auditivo (Alexander, 1962, p. 452).
De este modo, para Alexander, el origen de los cantos de los ortópteros se encuentra asociado al cortejo de este grupo de insectos. Esto es, el “primer canto” habría sido una señal de cortejo, cuya función es promover la asunción de la posición copulatoria en la hembra (Alexander, 1961, 1962). Las razones que posee Alexander para creer ello las vimos anteriormente. A partir de esta señal habría evolucionado nuestro canto de llamada, la cual, debido a ciertos cambios en la estructura y las propiedades auditivas del canto de cortejo (la intensidad, la duración y la ritmicidad) habría tenido un nuevo efecto ventajoso sobre los organismos, a saber, la atracción de hembras a distancias mayores, y este sería el origen de la función de atracción del canto de llamada. Esta sucesión de eventos evolutivos puede ser representada en un esquema como el siguiente:[1]
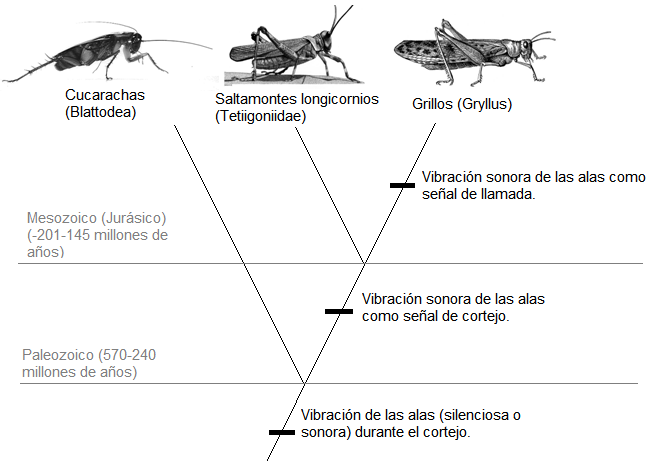
Figura 10: Cladograma esquemático de la historia filogenética de los cantos de llamada según Alexander.
Alexander, además, considera que la función secundaria de los cantos de llamada, esto es, el espaciamiento de los machos conespecíficos, habría “aparecido como una extensión de la función de llamada” (Alexander 1962, p. 453). La evidencia que menciona para creer ello puede resumirse como sigue:
- Las señales agresivas median las interacciones entre los machos. El canto de llamada también tiene tal función –y la diferencia es esencialmente cuantitativa. La señal de llamada sola provocará el repertorio agresivo completo de un macho agresivo.
- Las dos señales (las agresivas y las de llamada) difieren estructuralmente en una forma cuantitativa, y esto a su vez está relacionado en una forma cuantitativa con las diferencias funcionales. Esto es, cuanto más agresivo es el encuentro, más diferentes son los chirridos agresivos de los cantos de llamada. Pero incluso en las especies con las señales agresivas más distintivas, algunos de los chirridos agresivos son exactamente como los chirridos de llamada.
La evidencia es nuevamente comparada y funcional. (1) establece que los cantos de llamada también poseen la función de mediar las interacciones agresivas entre machos y (2) establece que las señales agresivas y de llamada se diferencian en mayor medida cuanto más agresivo es el encuentro macho-macho.
En base a esta evidencia, Alexander afirma que:
Cuando los cantos de llamada de los machos comenzaron a operar a larga distancia, parece inevitable que eventualmente desarrollaran importancia para los machos –los otros “tipos” de organismos de la especie- así como para las hembras. Primero, sabemos que la retroalimentación auditiva [auditory feedback] es uno de los primeros y más importantes atributos de la llamada en el macho –su habilidad de producir el canto con relativamente pocas interrupciones a través de largos periodos de tiempo. Un macho sordo raramente canta por largos periodos, y es cuestionable si la función de llamada en su requisito de redundancia podría haber precedido al desarrollo de órganos sensitivos auditivos en el macho. Segundo, hay ventajas selectivas obvias en las interacciones entre machos de cualquier animal en el cual los machos y las hembras se emparejan como resultado de interacciones comunicativas de largo alcance (Alexander, 1962, p. 455).
De este modo, para Alexander, la función secundaria del canto de llamada se habría desarrollado a partir de la función primaria, en tanto habría sido ventajoso para los machos (los cuales tienen la capacidad de oír sus propios cantos) reaccionar ante los cantos de llamada de otros machos de su especie (tal vez, en el contexto de la competencia por hembras).
Nótese que aquí las funciones son adquiridas evolutivamente en tanto resultan “ventajosas” para los organismos que poseen el rasgo con el efecto en cuestión (atraer hembras a larga distancia, provocar el espaciamiento de machos). En términos de Neander, dicha ventaja estaría determinada por la contribución al fitness inclusivo que realizaron tales efectos, y explican la presencia actual del rasgo en la especie. Aun aceptando esto, dado que los cantos tienen al menos dos funciones, y ambas habrían otorgado cierta “ventaja” a los organismos en el ambiente ancestral, no es posible establecer con precisión cuál de ambas es, en términos de Millikan, su “función propia”. Ambas funciones podrían explicar la presencia actual del rasgo en la especie. De hecho, el propio Alexander afirma que el canto de llamada habría tenido varios “roles” o “funciones”:
Este caso interesante [el de los gríllidos y tetigonideos] sugiere nuevamente el rol doble o múltiple del canto de llamada, y demuestra que una única señal puede ampliarse para involucrar más de una función en dos maneras obvias: a través de la misma señal teniendo diferentes efectos en diferentes situaciones o sobre distintos tipos de individuos, o a través del aislamiento de diferentes unidades funcionales en diferentes patrones componentes dentro del ritmo (en este caso, la llamada) (Alexander, 1962, p. 455).
Basándome en lo expuesto hasta aquí acerca de la evolución del canto de llamada según Alexander, creo que es posible afirmar dos cosas que perjudicarán la adecuación del enfoque etiológico para dar cuenta de las atribuciones funcionales de este caso de estudio.
En primer lugar, como uno puede sospechar de lo expuesto, las historias evolutivas, sobre todo las que refieren a rasgos comportamentales, suelen ser acusadas de ser altamente especulativas. Alexander mismo reconoce que…
Es difícil reconstruir los cambios evolutivos en el comportamiento desde el estudio comparativo solamente, y ciertos tipos de conclusiones deben siempre ser tentativas y especulativas (Alexander 1962, p. 443).
Sin embargo, las atribuciones funcionales no son objeto de dicha sospecha, y no parece necesario, desde el estudio de Alexander, establecer una historia evolutiva para establecer una atribución funcional. De hecho, la historia evolutiva podría haber sido reconstruida de una forma diferente, y aun así las atribuciones funcionales permanecerían idénticas.
En segundo lugar, como vimos, parte de la evidencia que utiliza Alexander para resolver la cuestión de la evolución del canto de llamada es la atribución de las funciones que establecimos previamente. Esto es, para comprender los caminos evolutivos que habría seguido el canto, primero atribuyó las funciones que conocemos. Alexander mismo considera que para entender los cambios evolutivos sucedidos en tales cantos es preciso conocer, primero, el tipo de funciones que cumplen dichos cantos:
Necesitamos primero entender las clases de señales, tanto estructural como funcionalmente, que los grillos poseen. Luego necesitamos tener alguna idea de la clase de acústico que fue el primer grillo (o algún grillo temprano), y necesitamos conocer cómo las diversas clases estructurales y funcionales de señales de los grillos están relacionadas, cómo nuevas funciones han evolucionado, y cómo la estructura ha cambiado para acomodarse a nuevas funciones (Alexander, 1962, p. 444).
Si la atribución funcional fuera equivalente a una afirmación acerca del origen evolutivo de los rasgos funcionales, como afirma el enfoque etiológico, dicha atribución funcional no podría ser evidencia de la evolución de dichos rasgos. Y, como acabamos de ver, la atribución funcional es parte de la evidencia que toma Alexander para establecer un posible camino evolutivo. Nótese la importancia de esta afirmación. Si la atribución funcional, que es evidencia para la evolución por selección natural, dependiera de la aplicación de dicha teoría, la teoría misma de la selección natural se tornaría circular. Esto es: si la identificación de funciones dependiera de la aplicación de la teoría de la selección natural, y a su vez dichas funciones fueran evidencia de la acción de la selección natural en el pasado, la teoría de la selección natural se autoconfirmaría en cada aplicación.
La carga de circularidad contra el enfoque etiológico no es una novedad de esta tesis. Dicha afirmación puede encontrarse, por ejemplo, en los trabajos de Ginnobili (2009, 2011) y de Caponi (2010).
La teoría de la selección natural utilizada por Darwin (que bajo mi punto de vista no difiere en este aspecto de la utilizada actualmente) requiere de un concepto de función independiente del origen histórico del rasgo, y particularmente, independiente de ella misma (Ginnobili, 2009, p. 6).
Puede afirmarse, por eso, que el gran malentendido que está en la base de la concepción etiológica del concepto de función es haber confundido este concepto con el de adaptación sin percibir que este es lógicamente posterior a aquel: primero tenemos que establecer la función de una estructura para después determinar si ella es o no una adaptación; es decir: si ella evolucionó o no por selección natural en virtud de las exigencias derivadas de ese desempeño funcional o en virtud de otras exigencias funcionales que también deberemos identificar. Las explicaciones seleccionales no justifican imputaciones funcionales, ellas las suponen y, a partir de otras consideraciones, nos permiten atribuirle a un rasgo el estatuto de adaptación. Es esta atribución, y no la atribución funcional, la que tiene un carácter etiológico (Caponi, 2011, p. 59).
Además de formar parte de la evidencia para la aplicación de la teoría de la selección natural, las atribuciones funcionales parecen constituir su explanandum. En efecto, como puede notarse en la explicación evolutiva de Alexander, el fenómeno del que se intenta dar cuenta es del origen de la función del canto de llamada (determinada previamente). Esto puede advertirse en fragmentos como el siguiente, ya citado anteriormente:
Eventualmente, mediante este tipo de cambio, este canto debió haberse vuelto operativo a tales distancias que a veces era ventajoso (esto es, en algunas instancias o en algunas especies) para el macho estridular sin contacto con la hembra, y a veces era ventajoso para la hembra ser atraída al escuchar esos sonidos cuando no estaba en contacto con el macho de otro modo. De este modo, la función de llamada, en la forma aproximada que asume hoy en día, podría haber evolucionado (Alexander, 1962, p. 452, mi énfasis).
Así, para Alexander, aquello que explica la evolución de la función de llamada es cierta ventaja pasada que habrían obtenido los machos al estridular en ausencia de las hembras, y que habrían obtenido las hembras al ser atraídas por los cantos así producidas en ausencia de otro contacto con el macho. Dicha ventaja, selección natural mediante, habría dado lugar a la presencia del rasgo funcional que conocemos hoy en día. De este modo, el explanandum de la explicación evolutiva es la adquisición del rasgo funcional presente en los organismos actuales. Pero, nótese nuevamente, esto requiere de la identificación previa del rasgo funcional de manera independiente de la teoría de la selección natural. De otro modo, la teoría de la selección natural se tornaría circular. Esto es, perdería explicatividad al identificar el fenómeno explanandum de manera dependiente de la teoría.
La idea de que la teoría de la selección natural explica la adquisición de funciones también puede encontrarse en los trabajos de Ginnobili (2009, 2013). Para él, la teoría de la selección natural explica lo que denomina “adecuación”, lo cual se caracteriza, en parte, como un rasgo que cumple una función efectivamente. Las adecuaciones, desde este punto de vista, son caracterizadas a partir de atributos como la complejidad (que sería difícilmente explicable como fruto de la casualidad), la coadaptación (esto es, el ajuste de partes entre sí) y, fundamentalmente, la presencia de funciones adecuadas a un fin. Aquí, como en nuestro caso, la explicación evolutiva daría cuenta de “cómo estas funciones han sido adquiridas a través de la historia evolutiva de la especie” (Ginnobili, 2009, p. 19).
Hasta aquí, el resultado de nuestra indagación apunta a la necesidad de elucidar el concepto de función de manera independiente de la teoría de la selección natural. Esto es, la teoría de la selección natural, si ha de ser explicativa, requiere de un concepto de función que pueda determinarse independientemente de ella misma (Ginnobili, 2009). En el siguiente apartado discutiré la atribución funcional a partir del caso estudiado, tomando en cuenta este requisito.
Si bien, según lo visto hasta aquí, obtuvimos evidencia de que los estudios evolutivos y los estudios funcionales son independientes y no deben ser confundidos, esto no implica que los estudios evolutivos no sean relevantes para los estudios funcionales. Esto es, si bien ambos tipos de estudios no se identifican, y no deben ser identificados, los estudios evolutivos pueden tener un rol heurístico para la búsqueda de funciones, y esta podría ser la intuición sobre la que se apoya el enfoque etiológico. Este rol heurístico puede evidenciarse al observar ciertas afirmaciones que realizaron los biólogos para este caso en particular.
En efecto, Alexander nota en diversas ocasiones (1957, 1960, 1967) que los cantos de llamada de diferentes especies de grillos, en especial aquellas que son similares y simpátricas, son diferentes estructuralmente. Dicho fenómeno, para Alexander, puede explicarse apelando a la selección natural, en especial, a la selección por el aislamiento reproductivo. Así, por ejemplo, Alexander (1967) afirma que:
Los eventos usuales del comportamiento sexual, en orden, son la formación de parejas, el cortejo, la copulación (o, mejor, inseminación) y, a veces, el mantenimiento de la pareja para inseminaciones adicionales. (…)
Aparte de la influencia de predadores, la acción selectiva sobre tales eventos puede ser vista únicamente como un asunto de maximizar la eficiencia de la secuencia dentro de la especie – esto es, disminuir el porcentaje de individuos sin pareja y el porcentaje de abortos prematuros de encuentros sexuales significativos (la cantidad de tiempo y energía perdidos). Pero, para cualquier especie, la naturaleza particular de las unidades individuales de la secuencia puede ser explicable sólo en el contexto del aislamiento reproductivo, en otras palabras, sólo en relación con la naturaleza de la misma unidad en el comportamiento de otras especies viviendo en la misma área y lo suficientemente parecidas como para ser potencialmente confusas para los animales mismos.
Si dos especies similares viven juntas el tiempo suficiente, parece probable que los eventos que ocurren tempranamente en la secuencia sexual con el tiempo se diferencien lo suficiente de tal manera que operen como un mecanismo de aislamiento reproductivo sin tener en cuenta la naturaleza de la diferencia original que hizo al entrecruzamiento o la interacción entre especies deletéreo. Entonces, las diferencias en la forma de formación de parejas prevendrían interacciones deletéreas entre especies similares más efectivamente (esto es, involucrando menos gasto de tiempo y energía) que, digamos, diferencias en el cortejo; las diferencias en el cortejo, a su vez, serían más eficientes que las barreras mecánicas para la copulación. Como resultado, la selección debe continuar para promover la divergencia en las señales de formación de parejas (o señales de agregación) (…) (Alexander, 1967, pp. 507-508, mi énfasis).
De este modo, sugiere Alexander, las diferencias en los cantos de llamada de diferentes especies, y en las señales de agregación en general, pueden deberse a (o haberse originado por) un mecanismo selectivo que promueve el aislamiento reproductivo de las especies, en tanto podría existir una ventaja en evitar la hibridación o el entrecruzamiento entre especies. El hecho de que los cantos de llamada de especies simpátricas sean diferentes, y la hipótesis de que dichas diferencias podrían haberse originado por un mecanismo selectivo, otorgan apoyo a la idea de que las hembras tendrían cierta capacidad de discriminar los cantos conespecíficos de los heteroespecíficos. Dicha sugerencia es tenida en cuenta, por ejemplo, en el siguiente pasaje:
El hecho de que no haya dos especies de insectos, no importa cuán cercanamente relacionadas, que se haya mostrado que tengan cantos de llamada o congregacionales inseparables, sugiere la probabilidad de una capacidad discriminatoria (Alexander, 1957, p. 110).
Esta sugerencia de Alexander, es explícitamente retomada por Walker (1957), quien a partir de ello supone (y muestra experimentalmente) que “los sonidos del macho de una especie atraen a las hembras de esa especie en particular y no de otras” (Walker, 1957, p. 626).
Estos experimentos muestran que los sonidos producidos por los insectos pueden ser importantes en el aislamiento reproductivo de las especies. Las diferencias menores en el ritmo o la tasa de los cantos pueden determinar si la hembra de una especie cantora responde a una canto o no. Si el canto es importante en el aislamiento de las especies, puede jugar un rol importante en la evolución de nuevas especies. Una vez que las diferencias en los cantos se desarrollaron y llegaron a ser significativas comportamentalmente, las especies hermanas productoras de sonido pueden vivir en contacto cercano con posibilidades reducidas de hibridación. Puede esperarse que los cantos de especies cercanamente relacionadas difieran en mayor medida que los caracteres que no influencian el mantenimiento del aislamiento de la especie (Walker, 1957, p. 634).
La idea de que las hembras podrían poseer la capacidad de discriminar entre los cantos de los machos de su especie y las de otras especies, y su demostración experimental, como vimos (sección 4.4), dio origen a una serie de estudios orientados a determinar qué parámetros del canto de llamada son los que resultan atractivos para las hembras y cómo sus sistemas nerviosos logran reconocer los parámetros del canto que son específicos de su especie.
En este punto, creo que lo que puede afirmarse es que la teoría de la selección natural operó como un factor heurístico que sugiere (y refuerza) ciertas hipótesis funcionales por sobre otras. En este caso, las diferencias entre los cantos de llamada de distintas especies, y el hecho de que exista una hipótesis evolutiva plausible sobre su origen, esto es, la hipótesis de la selección para el aislamiento reproductivo, operaría como un factor heurístico que sugeriría la presencia de una capacidad discriminatoria en las hembras −sugerencia, como vimos, sumamente fructífera−. Todo ello, además, refuerza la idea de que los cantos de llamada, y la fonotaxis de las hembras, funcionan como mecanismos de formación de parejas reproductivas. De hecho, la idea de que las diferencias en los cantos y las preferencias de las hembras habrían evolucionado por aislamiento reproductivo presupone que tales cantos y preferencias cumplen alguna función dentro de la reproducción de los insectos. De otro modo, no podrían tener un rol en el aislamiento reproductivo de las especies.
A pesar de, quizás, apoyarse sobre una intuición correcta, esto no debe ser considerado una concesión para el enfoque etiológico, puesto que, nótese nuevamente, las hipótesis funcionales no se identifican con las hipótesis evolutivas. Esto puede notarse al indagar la naturaleza de la hipótesis evolutiva de selección para el aislamiento reproductivo. La confirmación de dicha hipótesis depende, en parte, del hallazgo de evidencia de que existe un desplazamiento de caracteres en las zonas de simpatría (Searcy y Andersson, 1986), y dicha evidencia puede ser difícil de obtener por diversas razones (Walker, 1974; Searcy y Andersson, 1986). Más aun, puede ser difícil distinguir la hipótesis de evolución por el aislamiento reproductivo de la hipótesis de evolución por selección sexual, en tanto habría un solapamiento de las predicciones de ambas hipótesis, pudiendo ser incluso ambas verdaderas.[2] A pesar de que la evolución de las diferencias en los cantos y la capacidad discriminatoria de las hembras por aislamiento reproductivo es, de este modo, controversial, el hecho de que los cantos y la capacidad discriminatoria de las hembras tengan una función en la forma de reproducción de estos insectos no lo es. De hecho, nuevamente, la explicación evolutiva (la selección para el aislamiento reproductivo o la selección sexual) asume que los cantos y la capacidad discriminatoria de las hembras cumplen cierta función dentro del comportamiento sexual de estos insectos (la aproximación de los machos y las hembras). Y, nuevamente, asumir que dicha función es determinada por la historia evolutiva de estos rasgos, convertiría a la explicación selectiva en una explicación circular. En virtud de ello, lo único que puede afirmarse es que la teoría de la selección natural puede operar heurísticamente en la formación de hipótesis funcionales, pero las hipótesis funcionales son independientes de las hipótesis evolutivas.
Creo que este rol heurístico de la teoría de la selección natural puede ser visto más como un rol restrictivo sobre el tipo de fines que pueden perseguir los organismos que como un rol determinante sobre el tipo de fines que de hecho persiguen en cada caso particular. Esto es: la teoría de la selección natural podría imponer ciertas restricciones sobre las atribuciones funcionales, esto es, qué tipo de funciones pueden cumplir los rasgos de los organismos, pero no permiten determinar cuál es la función específica que cumple un rasgo particular de hecho. Y esto puede apreciarse si uno atiende al tipo de biología funcional que se realizaba de manera previa a la aparición de la teoría de la selección natural darwiniana.
Desde el punto de vista predarwiniano, las adaptaciones, de las cuales podía darse cuenta apelando a un diseñador o un creador, podían cumplir con fines orientados a sostener la economía natural o el mutualismo providencial (Caponi, 2011). Según estas ideas, los organismos no sólo presentan características que les permiten sobrevivir y reproducirse, sino que sus rasgos pueden responder a objetivos tales como el mantenimiento del equilibrio natural que, para los teólogos naturales, garantizaba la subsistencia de la creación toda. El mantenimiento de este equilibrio permitiría, por ejemplo, atribuir a los frutos la función de alimentar a los animales o a las plantas la función de purificar el ambiente (Paley, 1809). Más aun, el embellecimiento del mundo era un fin hacia el cual ciertos rasgos de los organismos podían estar orientados: según Paley (1809) tal sería el objetivo del color de la corola del tulipán y del plumaje adornado de las aves (para un tratamiento más profundo de estos y otros ejemplos, véase Ginnobili, 2013 y Caponi, 2011).
Sin embargo, desde el punto de vista darwiniano, los objetivos (o funciones) que pueden poseer los rasgos de los organismos se encuentran restringidos a aquello que puede reportarles un beneficio en términos de una mejora en su éxito reproductivo diferencial (Ginnobili, 2013), en tanto tales son el tipo de funciones cuyo origen puede ser explicado por la teoría de la selección natural. Tales objetivos pueden ser la nutrición, la evitación de predadores, la reproducción, etc. En cambio, los rasgos cuyos efectos parecen estar orientados al beneficio de otra especie, al mantenimiento del equilibrio natural, o al embellecimiento del mundo, no pueden ser explicados desde un punto de vista darwiniano, en tanto no son el tipo de adaptaciones que pueden ser mantenidas con la lógica de la selección natural. En este sentido es que creo que podría decirse que la teoría de la selección natural restringe el tipo de objetivos que pueden satisfacer los rasgos de los organismos, esto es, aquellos que pueden mejorar su éxito reproductivo diferencial. Sin embargo, este rol restrictivo no es, como dije, un rol determinante. La teoría no determina qué objetivo satisface un rasgo en particular, o qué función cumple.
Aplicando esta distinción al caso que nos ocupa, la teoría de la selección natural podría haber cumplido un rol restrictivo sobre el tipo de roles biológicos que pueden cumplir los rasgos biológicos, a saber, la atracción de hembras, o el hallazgo de machos conespecíficos, en tanto tales son el tipo de roles que pueden ser favorecidos por un mecanismo selectivo (sea este el aislamiento reproductivo, o la selección sexual). Sin embargo, esta restricción sobre los roles o funciones que pueden cumplir los rasgos no debe ser confundida con la determinación efectiva de tales funciones biológicas, ni con la evidencia, utilizada por los biólogos, para realizar esta determinación. En consecuencia, en el próximo apartado abordaré la cuestión de cómo pueden determinarse las funciones biológicas de manera independiente de la selección natural.
5.1.2. La determinación de la función biológica
Un modo de abordar la cuestión sobre el modo en el cual los científicos determinan las funciones de los rasgos biológicos es observar el tipo de evidencia que utilizan para realizar dicha determinación. En consecuencia, en este apartado abordaré la cuestión de la determinación de las funciones biológicas enfocándome en el tipo de evidencia que utilizaron los biólogos para atribuir a los cantos de llamada la función primaria de atraer hembras conespecíficas.
Tal como expuse en el capítulo anterior, una parte de la evidencia proviene de las observaciones de Alexander sobre el comportamiento de los grillos y su relación con otros comportamientos presentes en estos insectos. Tales observaciones son representadas por Alexander en lo que, en términos de la etología clásica, se denominaría un etograma. Los etogramas, en el sentido clásico (Tinbergen 1989 [1951]), constituyen una descripción del repertorio de comportamientos observados en una especie, en términos de posturas corporales, patrones de movimientos más o menos estereotipados, producción de sonido, cambios de color, descargas eléctricas, etc. (Zupanc, 2010). En este caso, como puede apreciarse en los trabajos de Alexander (1957, 1960, 1961, 1967) el repertorio de comportamientos observados en los grillos es descrito y representado una y otra vez mediante descripciones y gráficos como el de las figuras 3 y 4 (capítulo 4). En estos gráficos, la producción de sonidos por parte de los machos causa la atracción de las hembras hacia los machos. Si uno continúa las secuencias indicada por los gráficos, todo ello decanta en la copulación de los sexos y la ovoposición. En virtud de ello, Alexander mismo indica, como ya hemos visto (sección 4.3), que la función del canto de llamada es la atracción de hembras conespecíficas, en tanto es aquello que contribuye a la copulación de estos organismos.
Más aun, Alexander (1997) inserta esta secuencia de comportamientos dentro de lo que describe como los componentes de “la secuencia de apareamiento” [mating sequence] o “secuencia reproductiva” [reproductive sequence]. Tal secuencia estaría compuesta por ocho componentes (no siempre presentes en todas las especies): (1) acercamiento o formación de parejas, (2) cortejo, (3) copulación, (4) inseminación, (5) eventos post-copulatorios o intercopulatorios, (6) fertilización, (7) cuidado parental cooperativo y (8) unión a largo plazo. Según Alexander (1997: 6) la fase (1) puede involucrar (a) miembros de uno o ambos sexos buscando activamente al otro en el hábitat apropiado, (b) miembros de ambos sexos encontrándose en algún lugar como resultado de un estímulo extrínseco tal como el olor de las plantas de ovoposición o patrones visuales del ambiente o (c) miembros de un sexo señalando (“llamando”) a miembros de otro sexo.
Dentro de esta secuencia, el canto de llamada se inserta como parte de (1) el acercamiento o la formación de parejas, en tanto uno de sus efectos (el que se considera su función) consiste en (c) atraer (o “llamar”) hembras conespecíficas. De este modo, creo que, en parte, lo que permite atribuir esta función al canto de llamada es la consideración de que contribuye a la capacidad de los grillos, y de otros insectos, de reproducirse (y por ello forma parte de la secuencia reproductiva). En este sentido, la representación de la secuencia de comportamientos que exhibe el etograma permitiría al etólogo determinar cuál es la función de cada componente, teniendo en cuenta ciertos objetivos razonables, desde el punto de vista darwiniano, al cual pueden contribuir. En este punto, esto puede ser pensado como un análisis funcional a la Cummins (sección 3.2) pero procediendo de manera bottom-up (de abajo hacia arriba). Esto es: en lugar de pensar cómo puede implementarse una capacidad de manera top-down (de arriba hacia abajo), descomponiendo dicha capacidad en otras sub-capacidades y sub-componentes, aquí se estaría procediendo pensando a qué capacidad contribuye toda esta secuencia de comportamientos que tenga sentido desde un punto de vista darwiniano, y atribuyendo funciones a cada evento de la secuencia en virtud de su contribución a dicha capacidad (en este caso, reproductiva). Poder brindar una visión acabada y completa del modo en el que todas las partes contribuyen al cumplimiento de la función o capacidad, parece servir de evidencia en favor de las atribuciones funcionales a cada una de estas partes. En el apartado siguiente volveré sobre la aplicación del enfoque sistémico a este caso.
En su intento de comprender el rol que cumplen los distintos sonidos de los artrópodos e insectos en general (Alexander, 1957, 1967) y de los ortópteros en particular (Alexander, 1960), Alexander ofrece una gran cantidad de comparaciones en torno a los efectos que producen estos sonidos en distintas clases de insectos. En este contexto, otro tipo de evidencia utilizada por Alexander para el establecimiento de la función del canto de llamada en los grillos es la evidencia proveniente de la comparación de los efectos de aproximación que posee este comportamiento (y otros) en los insectos. Esta evidencia podría subdividirse en dos tipos. Por un lado, Alexander ofrece comparaciones entre insectos en los cuales la producción de sonidos con un efecto de aproximación es presumiblemente un rasgo homólogo (compartido debido a que poseen un ancestro común), entre los tetigonideos (saltamontes) y los gríllidos (grillos) (Orthoptera) (Alexander, 1957, 1960, 1967). En estas comparaciones puede notarse que en la atribución de funciones no sólo tiene relevancia la selección natural (en la que los defensores del enfoque etiológico se han enfocado únicamente), sino también el estudio de rasgos homólogos. Esto es: el hecho de que especies cercanas usen un rasgo con cierto objetivo (la atracción de hembras), podría ser evidencia de que la especie bajo estudio lo usa de la misma manera.
Sin embargo, por otro lado, Alexander también tiene en cuenta el rol de estos sonidos en animales en los cuales estos sonidos no constituyen rasgos homólogos, sino análogos (por ejemplo, en los acrídidos (langostas), los cicádidos (cigarras) y en ciertos crustáceos) (Alexander, 1967). Asimismo, Alexander considera otros grupos de insectos en los cuales los sonidos que causan la aproximación de los sexos son ligeramente diferentes, en tanto son ejecutados por las hembras (Culcidae ─mosquitos─, Ceratopogonidae ─chinches─, Trogidae ─escarabajos─) o por hembras y machos en alternancia (Acridinae ─saltamontes de campo─, Tettigoniidae ─esperanzas de alas angulares─, Curculionidae ─gorgojo de la ciruela─). Más aún, Alexander menciona evidencia proveniente de otros tipos de señales que tienen el mismo efecto de aproximación en otros grupos de insectos: estímulos odoríferos en los lepidópteros (mariposas), estímulos visuales (patrones de color, patrones de movimiento en el vuelo) en ciertos dípteros (moscas), ritmos de intermitencia en la luz emitida por ciertos coleopteros (luciérnagas), etc. (Alexander, 1967). Nótese que en estos casos la evidencia comparada es heterogénea, entre especies de insectos de distintos ordenes, y con respecto a rasgos que no son similares (patrones de color, intermitencias de luz, etc.) sino sólo con respecto a su efecto de aproximación de las parejas sexuales. Este tipo de evidencia podría interpretarse como evidencia de la existencia de este tipo de señales de aproximación (o funciones) en la naturaleza. Tal como Alexander lo expresa:
La función ha sido demostrada más frecuentemente para la formación de parejas o las señales de llamada que para cualquier otra clase de señal acústica de los insectos. Más a menudo, la formación de parejas se logra por la simple atracción de las hembras sexualmente receptivas hacia la señalización masculina (Alexander, 1967, p. 508).
Este tipo de evidencia comparada no es tenida en cuenta por los enfoques que presenté en el capítulo 3, con la excepción de Horan (1987), quien la considera relevante para la formación de lo que denomina “hipótesis funcionales”. Según la autora, la evidencia comparada es de dos tipos: (i) evidencia comparada de rasgos similares (presumiblemente homólogos) de especies cercanamente relacionadas, y (ii) evidencia comparada de rasgos similares (presumiblemente homoplasias) de especies distantes o no relacionadas. Estos dos tipos de evidencia darían lugar, según Horan, a distintos tipos de inferencias. Evidencia del tipo (i) permitiría al investigador relacionar las diferencias entre los rasgos homólogos con ciertos correlatos ecológicos (o presumibles presiones selectivas) que darían cuenta de tales diferencias, mientras que la evidencia de tipo (ii) permitiría al investigador relacionar las similitudes entre los rasgos análogos con ciertos correlatos ecológicos que darían cuenta de tales similitudes. Ambos tipos de evidencia permitirían al investigador, en una fase que Horan llama “inductiva”, formular una “hipótesis funcional” que explique la correlación entre el rasgo en cuestión y su factor ecológico asociado, en tanto éste último constituiría aquella “presión selectiva” frente a la cual el rasgo tendría un efecto que aumenta el fitness de los organismos que lo poseen. Es decir, la apelación a la evidencia comparada es considerada por Horan como funcionando en el contexto de descubrimiento de la atribución funcional, la cual debe ser confirmada al mostrarse que el efecto funcional contribuye al fitness de los organismos portadores. Sin embargo, lo que parece ser el caso aquí, es que la atribución funcional ocurre, en primer lugar, sin haberse identificado una presión selectiva o un factor ecológico que explique las similitudes o las diferencias entre los rasgos, y, en segundo lugar, sin haberse indagado la contribución al fitness que reporta su posesión para los organismos (volveré sobre este punto más adelante). Lo que este tipo de evidencia comparada parece mostrar, antes bien, es que habría un efecto compartido por todos estos rasgos (y esta es su única similitud): la atracción de hembras, o la formación de parejas. Y nótese que dicho efecto no sólo es importante por ser ubicuo. Muchos de estos rasgos también poseen el efecto de atraer predadores, pero tal no es el efecto que se destaca en la atribución funcional. Parte de la importancia que poseen, sugiero, es que contribuyen a una capacidad darwinianamente importante de los organismos: su reproducción.
Sin embargo, esta no es la única evidencia utilizada para determinar la atribución funcional. También se realizaron ciertos experimentos (sección 4.4.1) orientados a determinar si es el canto de llamada de los grillos aquello que atrae a las hembras (y no otra cosa), y cuáles son los parámetros del sonido que son los relevantes para causar dicha respuesta fonotáctica. De este modo, en primer lugar, se encuentra la contrastación experimental de Regen (1913), quien mostró que las hembras de grillo podían ser atraídas por el canto de un macho transmitida a través de un receptor telefónico, aislando, de este modo, otras variables que podrían influir en su respuesta, tales como estímulos visuales, táctiles o químicos. De un modo un poco más sofisticado, Walker (1957) logró mostrar que los cantos de los machos atraen a hembras de la misma especie, y no de otras especies, notando incluso que las diferencias entre los cantos radicaban principalmente en la tasa de sílabas por segundo que poseían. Posteriormente, surgieron una gran cantidad de experimentos orientados a determinar qué patrones del sonido son aquellos que la hembra detecta y le permiten reconocer y localizar a los machos de su especie, coincidiendo en que la tasa de sílabas por segundo es uno de los patrones más relevantes para el reconocimiento en los grillos de campo (sección 4.4.1). Sin embargo, nótese, estos experimentos sólo exhiben una relación existente entre el rasgo (el canto de llamada o alguna de sus propiedades) y uno de sus efectos (la atracción de hembras). Esto es: estos experimentos muestran que el rasgo es condición suficiente para el efecto, pero no que dicho efecto sea la función del rasgo. De hecho, se ha mostrado experimentalmente que los cantos de llamada son condición suficiente también para, por ejemplo, la atracción de ciertos predadores o parásitos (Walker, 1964; Cade, 1975; Bell, 1979; Sakaluk y Belwood, 1984) pero no se dice que tal efecto sea su función. Por ello, la demostración experimental de que el canto es condición suficiente para la producción de cierto efecto no es en sí mismo suficiente para afirmar que tal efecto es su función.
Otros experimentos se han orientado en el sentido opuesto, intentando mostrar si el rasgo es condición necesaria para el efecto. En esta dirección se halla, por ejemplo, el experimento realizado por Duijm y van Oyen (1948), y mencionado por Walker (1957), quienes mostraron que las hembras de chicharra alicorta (Ephippiger ephippiger) son atraídas por machos cantores ocultos, pero no por machos que han sido silenciados mediante el pegado de sus alas. De este modo, se infiere que el movimiento de las alas (necesario para la estridulación y el canto) es condición necesaria para la atracción de hembras. Sin embargo, nuevamente, mostrar que un rasgo es condición necesaria para un efecto no es suficiente para mostrar que ese efecto es la función del rasgo, en tanto, nuevamente, uno podría mostrar que los cantos de llamada son condición necesaria para la atracción de predadores o parásitos y aun así no afirmaría que tal es la función del canto.
A pesar de que este tipo de evidencia parece mostrar que el canto de llamada es condición necesaria para la atracción de parejas (al menos en la chicharra alicorta), no es ésta la única forma en la que los grillos logran hallar una hembra de su especie. En efecto, se han encontrado machos no-cantores diversas especies de grillos: G. integer (Cade, 1979, 1984), G. Rubens (Burk, 1982), T. oceanicus (Zuk, Simmons y Cupp, 1993) y G. campestris (Hissmann, 1990). Si bien no parece haber acuerdo respecto de por qué ciertos machos no cantan (véase las revisiones de Greenfield, 1997; Zuk & Simmons, 1997), algunos investigadores creen que tales machos hallan parejas interceptando a las hembras atraídas por los machos cantores (Cade 1979, 1984), adoptando una estrategia de apareamiento “satélite” y evitando así el riesgo de predación y parasitismo. De hecho, un mismo individuo puede alternar entre cantar y no cantar dependiendo de la densidad de la población en la que se encuentra inmerso (Alexander, 1961; Cade y Cade, 1992; Rowell, Cade y Unicersity, 1993), obteniendo parejas o bien cantando o bien caminando por el territorio sin cantar. De este modo, lo que resulta de estos hallazgos es que el canto de llamada no es estrictamente condición necesaria para la formación de parejas (aunque puede serlo para la atracción de hembras), en tanto existiría una estrategia conductual alternativa que conduciría al mismo fin (ser un macho satélite). Lo que esto muestra es que, para la determinación de la atribución funcional del canto de llamada, no se requiere que el rasgo sea condición necesaria para la consecución del objetivo en virtud del cual se le atribuye dicha función. Esto es: no se requiere que el canto sea condición necesaria para la formación de parejas o la reproducción, en tanto existen otras formas de conseguir dicho objetivo (por ejemplo, siendo un macho satélite).
Dentro de la discusión filosófica sobre el concepto de función en biología, esto ha sido denominado el problema de los equivalentes funcionales, señalado inicialmente por Hempel (1979, sec. La lógica del análisis funcional). Para Hempel, el hecho de que existan diversas estructuras que pueden cumplir una misma función (los equivalentes funcionales), esto es, que ninguna sea condición necesaria para el cumplimiento de la función, tiene como consecuencia que uno no puede inferir la presencia de ninguna de las estructuras a partir del hallazgo de que una función se cumple y, por lo tanto, tampoco puede explicar la presencia de ninguna de tales estructuras según su modelo inferencialista de la explicación.
En virtud de lo visto, es posible decir que para la determinación de la atribución funcional no es necesario ni suficiente establecer que el rasgo es condición necesaria y/o suficiente para la producción del efecto que se determina como su función. Por un lado, mostrar que el rasgo es condición necesaria para su efecto funcional no es condición necesaria ni suficiente para la atribución funcional porque, debido al problema de los equivalentes funcionales, puede ocurrir que un rasgo no sea condición necesaria para el cumplimiento de su función, y aun así se establezca que cierto efecto es su función. Este es el caso, por ejemplo, del canto de llamada y su equivalente funcional: el comportamiento satélite. Por otro lado, mostrar que el rasgo es condición suficiente para su efecto funcional no es condición necesaria ni suficiente para la atribución funcional, debido a que mostrar que un rasgo es condición suficiente para un efecto no siempre da lugar a una atribución funcional. Este es el caso, por ejemplo, de las demostraciones experimentales orientadas a mostrar que los cantos de llamada son suficientes para la atracción de predadores y parásitos. Por supuesto, esto no implica que tales estudios no sean relevantes. Evidentemente juegan un rol fundamental para la atribución funcional (y los investigadores lo juzgan así). Más adelante volveré sobre esta cuestión de la naturaleza de la atribución funcional, en ausencia de criterios necesarios y suficientes para la aplicación del concepto de función.
En la dirección de establecer si el rasgo es condición necesaria para el efecto, también se encuentra el razonamiento contrafáctico que, en parte, estableció que las hembras deberían poseer la capacidad de discriminar los cantos conespecíficos de las heterospecíficas. Dicho razonamiento es realizado por Alexander y Walker cuando piensan qué pasaría si la hembra se atrajera por todos los cantos que escucha (si no distinguiera los cantos conespecíficos de las heterospecíficas).
(…) la incapacidad de las hembras para reconocer los cantos de los machos de su propia especie resultaría en una confusión total (…) Si esa hembra respondiera igualmente a todos los sonidos de los insectos de la zona, o incluso al sonido más intenso en su vecindad, su posibilidad de conocer a un macho de la misma especie se reduciría mucho y el tiempo requerido haría que esto fuera un sistema de comunicación ineficiente (Alexander, 1957, p. 111, mi énfasis).
Muchas especies diferentes cantan al mismo tiempo en el mismo hábitat. Si una hembra respondiera indiscriminadamente a tales sonidos, ella podría ir hacia diversos machos de cada una de muchas especies antes de toparse con uno de su propia especie. Esto tendría un valor de supervivencia dudoso (Walker, 1957, p. 626).
Dicho razonamiento contrafáctico establece que, si las hembras no discriminaran los cantos propios de su especie, no hallarían a un macho conespecífico (reduciendo sus posibilidades de reproducción). En consecuencia, se considera que la posesión de una capacidad discriminatoria en las hembras es condición necesaria para el hallazgo de machos conespecíficos y, por ende, la formación de parejas reproductivas. Entonces, si, para la formación de parejas, la función de los cantos de llamada es la atracción de hembras (conespecíficas), la función de la (supuesta) capacidad discriminatoria de las hembras es el hallazgo de machos conespecíficos. Si dicha capacidad discriminatoria no estuviera presente (contrafáctico), entonces las hembras tendrían grandes dificultades en hallar un macho de su especie. Sin embargo, nótese, inferir que la hembra debe poseer una capacidad discriminatoria a partir de un razonamiento contrafáctico no implica qué tipo de estructura (neural) debe subyacer a dicha capacidad. Y esto sucede en virtud del problema de los equivalentes funcionales: conocer que una función se cumple no implica qué estructura debe estar presente para cumplirla. Esto es, precisamente, la tarea abordada por la indagación neuroetológica que veremos en la próxima sección.
Este tipo de razonamientos o comparaciones contrafácticas en las atribuciones funcionales son considerados fundamentales de la biología funcional dentro del enfoque de valor de supervivencia (sección 3.3) en autores como Canfield (1964) y Bigelow y Pargetter (1987). Wouters (1999) también considera tales comparaciones como una fuente importante de evidencia en la biología funcional. De manera general, según estos autores, la comparación de la situación actual (las hembras logran distinguir –y encontrar– a los machos conespecíficos a partir de sus cantos de llamada) con una situación contrafáctica (las hembras son incapaces de distinguir los cantos de llamada específicas de su especie de las heterospecíficas) en la que es, de algún modo, evidente que las probabilidades de supervivencia o reproducción de los organismos se verían disminuidas, es una fuente de evidencia para la atribución funcional (la capacidad discriminatoria de las hembras tiene la función de distinguir a los machos conespecíficos). De este punto dos cosas resultan interesantes.
En primer lugar, aquello a lo que se le atribuye una función (la capacidad discriminatoria) es algo supuesto: aún se desconocía el mecanismo neural que cumplía esa función y, como vimos, fueron necesarios ciertos hallazgos neurofisiológicos para confirmar la existencia de dicha capacidad de las hembras, en términos estructurales. Por lo que aquí el razonamiento contrafáctico estaría haciendo algo más que determinar una función: estaría determinando la presencia de un mecanismo subyacente que satisfaga dicha función (aunque se desconozca su estructura). La búsqueda de un mecanismo neural que cumpla dicha función, y el lenguaje funcional utilizado en dicha indagación, serán cuestiones a tratar en la próxima sección.
En segundo lugar, es interesante notar que, dentro del enfoque de valor de supervivencia (véase la sección 3.3), la contribución a las probabilidades de supervivencia y reproducción es aquello que determina si un efecto de un ítem o rasgo es funcional o no. La elección de este tipo de objetivo último dentro de la biología funcional, como vimos en la sección anterior, tiene sentido desde un punto de vista darwiniano, pero no dentro de puntos de vista como la teología natural, en donde los objetivos últimos de la naturaleza podían incluir el equilibrio natural, el mutualismo providencial o el embellecimiento del mundo. Por lo que, en este punto, creo que un aporte fructífero de este enfoque reside en determinar un objetivo último que pueden perseguir los rasgos de los organismos de manera consistente con el rol restrictivo de la teoría de la selección natural, pero sin proponer a la teoría de la selección natural como un factor determinante de las funciones que cumplen los ítems o rasgos en cada caso particular. Además, parte de la evidencia que se utiliza en dichas atribuciones, como acabamos de ver, incluye la formulación de experimentos o razonamientos contrafácticos del tipo propuesto por este enfoque.
A pesar de que el enfoque de valor de supervivencia logra dar cuenta de cierto tipo de evidencia utilizada para determinar las atribuciones funcionales (evidencia contrafáctica y comparada), la sugerencia realizada por Horan (1987) (cf. sección 3.3) de que la determinación de una función próxima (o la confirmación de una hipótesis funcional) depende de mostrar cómo el efecto funcional contribuye al fitness del organismo que lo posee (y así justificar la atribución de una función última) parece inadecuada. En el contexto de esta investigación, la atribución de la función próxima consistiría en afirmar lo siguiente: “la función próxima del canto de llamada es atraer hembras conespecíficas”. Mientras que la atribución de la función última consistiría en afirmar que “la función última del canto de llamada es aumentar el fitness de los machos que la poseen”.
Según Horan, para la atribución de una función próxima a un rasgo uno debe mostrar cómo dicho efecto aumenta el fitness del organismo que lo posee. Una forma de comprobar la adecuación del enfoque de Horan reside en verificar si los biólogos realizaron una medición de fitness a los fines de atribuir esta función próxima. Nótese que esta prueba de adecuación también se aplicaría a la postura de Walsh (1996) y de Kitcher (1993) dentro del enfoque de selección no histórica (cf. sección 3.5), en tanto, desde estos puntos de vista, la función de los rasgos se establece en relación a su contribución al fitness del organismo dentro de cierto régimen selectivo (en este caso, determinado por el ambiente actual) (Walsh, 1996) o bien en el pasado reciente y el presente (Kitcher, 1993). Como vimos, dentro de las perspectivas de Horan, Kitcher y Walsh, el fitness es considerado una propensión de los organismos a sobrevivir y reproducirse, y las funciones de los rasgos son determinadas cuando se muestra que el efecto funcional aumenta dicha propensión de los organismos poseedores del rasgo. Sin embargo, “nadie mide ‘propensiones’ de los organismos a hacer cosas” (Hunt & Hodgson, 2010, p. 47), por lo que, en la práctica (en este caso, de la ecología del comportamiento) para medir el aporte que realiza cierto rasgo al fitness de los organismos, suele medirse alguna variable que se considera que correlaciona positivamente con el fitness, tales como el éxito reproductivo o algún proxy del fitness (la atractividad de un macho, la cantidad de energía consumida, la calidad del territorio controlado, etc.).
En el contexto de la investigación sobre los cantos de llamada de los gríllidos, este tipo de medición fue realizada utilizando el “éxito en el apareamiento” [mating success] como un proxy del fitness[3], con el fin de establecer la ventaja que reporta (en términos de fitness) la adopción de las siguientes “estrategias reproductivas” bajo ciertas condiciones ambientales: ser un macho cantor (“llamar” hembras desde la madriguera) o ser un macho no-cantor (permanecer en la madriguera o recorrer cierto territorio sin cantar, esto es, ser un macho “satélite”). De manera general, estos estudios intentan establecer correlaciones entre ciertas condiciones ambientales (la densidad de la población, la proporción de sexos, la cantidad de machos adoptando la estrategia reproductiva alternativa, etc.) y el “éxito en el apareamiento” que reporta para los individuos adoptar alguna de estas estrategias. Así se halló, por ejemplo, en G. integer, que a altas densidades de población, el éxito en el apareamiento es mayor para los machos no-cantores, mientras que, a bajas densidades, los machos cantores son más exitosos (Rowell, Cade y Unicersity, 1993). En G. Campestris se mostró que, a altas densidades de población, los machos no-cantores fueron más exitosos en el hallazgo de hembras conespecíficas (medido en número de encuentros), mientras que, a bajas densidades, los machos cantores fueron más exitosos. Sin embargo, tanto a altas como bajas densidades de población, los machos cantores fueron más exitosos en términos de apareamientos logrados (posiblemente, debido a que los machos no-cantores reaccionaban agresivamente en muchos de los encuentros sexuales logrados) (Hissmann, 1990). La cantidad y complejidad de estudios orientados en esta dirección, la discrepancia de los resultados, y su variedad, dependiendo de la especie estudiada, el método elegido y el objetivo concreto de cada investigación, hace difícil establecer una respuesta concreta respecto de la contribución al fitness que reporta la adopción de la estrategia de los machos cantores en cada situación particular. Lo que parece ser el caso es que, bajo ciertas condiciones ambientales, la estrategia de los machos cantores es ventajosa (contribuye al fitness en mayor medida que la estrategia alternativa), mientras que, bajo ciertas otras condiciones ambientales, la estrategia puede ser desventajosa, o contribuir en menor medida al fitness que la estrategia alternativa.
Si este es el caso, entonces es posible dar cuenta de por qué las posturas de Horan (1987), de Walsh (1996) y de Kitcher (1993) resultan inadecuadas. Según estos autores, un rasgo (el canto de llamada) tiene una función (atraer hembras) en cierto contexto (cierta densidad de población) si contribuye al fitness de los organismos que lo poseen (en relación con la contribución de un rasgo alternativo). Sin embargo, la atribución de esta función al canto de llamada no parece ser dependiente de si contribuye al fitness de los organismos en determinado contexto. Como vimos, en ciertos contextos es una estrategia ventajosa, y en ciertos otros contextos no lo es (o no es la más ventajosa), pero, en todos los contextos, es una estrategia reproductiva, en tanto posee la función de atraer hembras. Es decir: considerar que el canto de llamada es parte de una estrategia reproductiva es un presupuesto de estos estudios orientados a comparar la contribución al fitness de esta estrategia frente a otra. Y considerar que es parte de una estrategia reproductiva es considerar que tiene una función en la reproducción de estos organismos, tanto cuando es ventajosa como cuando no lo es. Lo mismo sucede con la estrategia alternativa. Los machos no-cantores, se considera, adoptan la estrategia satélite, y dicha estrategia es ventajosa en ciertos contextos, pero no en ciertos otros. Pero no deja de ser una estrategia reproductiva cuando no es ventajosa, esto es, no deja de tener una función reproductiva cuando los individuos que la adoptan se encuentran en desventaja frente a la estrategia alternativa. De este modo, lo que parece ser el caso, es que la atribución de una función al canto de llamada no es dependiente de determinar si dicho efecto contribuye al fitness de los organismos en determinado contexto. Al contrario, presuponer que tiene dicha función es parte de considerarla una estrategia reproductiva cuya contribución al fitness puede variar dependiendo del contexto en cuestión.
En definitiva, la evidencia mostrada hasta aquí para la atribución funcional en cuestión podría resumirse en la siguiente lista: (i) la realización de un etograma y la estrategia de razonamiento bottom-up (¿a qué capacidad ─darwiniana─ contribuye esta serie de comportamientos con estos efectos?), (ii) la evidencia comparada de efectos similares de comportamientos de organismos emparentados y no emparentados, (iii) la realización de experimentos que muestren si el comportamiento es condición necesaria y/o suficiente para el efecto y (iv) la formulación de razonamientos contrafácticos (¿qué sucedería si no se diera cierto efecto?). Como vimos, algunos enfoques filosóficos pueden dar cuenta de la forma en la que se utiliza parte de esta evidencia. Así, el enfoque sistémico podría dar cuenta, por ejemplo, de (i), en tanto la estrategia de razonamiento bottom-up procedería a la manera de un análisis funcional pero en sentido inverso, insertando el comportamiento en cuestión (el canto de llamada) dentro de una secuencia de comportamientos (en términos de Alexander, la secuencia reproductiva) cuya manifestación ordenada contribuye, como un todo, a la capacidad del organismo de reproducirse. Por otra parte, vimos que, dentro del enfoque de valor de supervivencia, podía darse cuenta del uso de la evidencia comparada (ii) y de la formulación de razonamientos contrafácticos (iv) para establecer la función de los rasgos, en tanto permitirían destacar ciertos efectos en virtud de su contribución a una capacidad que se considera importante para los organismos desde un punto de vista darwiniano.
Sin embargo, como vimos en el capítulo 3, los distintos enfoques filosóficos pretenden ofrecer una definición del concepto de función mediante el establecimiento de ciertas condiciones necesarias y suficientes para su aplicación. En este sentido, la evidencia precedente es, según cada enfoque, una condición necesaria para la aplicación del concepto de función. Pero, como vimos antes en el caso de la evidencia (iii), ninguno de estos tipos de evidencia define condiciones necesarias y/o suficientes para la aplicación del concepto. En particular, la evidencia ofrecida por la aplicación de las estrategias (i), (ii) y (iv) dependen de la consideración de que hay una capacidad relevante de los organismos a la cual el efecto funcional contribuye, pero dicha capacidad relevante no está definida previamente a la aplicación de la estrategia. El canto de llamada pertenece a una secuencia reproductiva en tanto uno de sus efectos provoca la aproximación de las hembras y permite la formación de parejas reproductivas. Pero no forma parte de dicha secuencia reproductiva previamente a establecer que su función es la formación de parejas reproductivas. Por lo que, lo que parece ser el caso, es que la capacidad relevante a la que contribuye el efecto funcional se establece junto con, y no previamente a, la atribución funcional. Nótese que, obviamente, no siempre la reproducción es la capacidad relevante. En otros casos, la capacidad relevante puede ser la evasión de predadores, la detección de presas, el cuidado adecuado de las crías, etc. En este sentido, si la capacidad relevante no se encuentra definida previamente, es difícil ver cómo esta evidencia puede constituir un conjunto de condiciones necesarias y/o suficientes para la aplicación del concepto de función, y de este modo definirlo. Antes bien, este tipo de evidencia se asemeja más al tipo de criterios de determinación que son característicos de los términos teóricos en el sentido de Hempel (1979). Según Hempel, los términos teóricos poseen la particularidad de no ser definibles a partir de un conjunto de condiciones necesarias y suficientes formuladas en lenguaje observacional.
Cualesquiera que sean los criterios observacionales de aplicación que el científico pueda proporcionar, lo que tienen por objeto es simplemente describir síntomas o indicaciones de la presencia de la entidad en cuestión y no dar una caracterización exhaustiva de ella (Hempel, 1979, p. 209).
Entender el concepto de función como un concepto teórico permitiría entender a las condiciones (i)-(iv) como síntomas o criterios de determinación del concepto, en lugar de entenderlas como condiciones necesarias y/o suficientes para su definición. Y comprenderlas de esta manera tal vez permita enfocar de un modo diferente el problema de la elucidación del lenguaje funcional. Desde este punto de vista, la elucidación del concepto de función no consistiría en hallar una definición (que para los términos teóricos no es posible), sino en hallar una teoría subyacente al concepto junto a sus criterios de determinación. El intento de hallar una teoría subyacente ya fue realizado en algunas ocasiones (Ginnobili, 2009, 2011), y en trabajos posteriores podría ser fructífero continuar con este punto de vista. Aquí sólo dejaré establecido que los criterios precedentes podrían ser tomados fructíferamente como síntomas para la aplicación del concepto teórico de función, que de aquí en adelante mencionaré como función biológica.
Recapitulando lo visto acerca de determinación de la función biológica en el caso que nos ocupa, uno puede encontrar que los científicos apelaron a un conjunto heterogéneo de evidencia, evolutiva y no evolutiva. Los diferentes enfoques han intentado dar cuenta del concepto de función sólo ocupándose de alguno de estos tipos de evidencia, y ensayando una definición explicita del concepto de función en base a ella. Lo que he intentado mostrar, a partir del análisis del caso visto, es que, si bien todos estos tipos de evidencia de hecho han cumplido un rol en la atribución funcional, ninguna de ellas por sí misma, ni todas ellas en conjunto, brindan criterios de condiciones necesarias y suficientes de la atribución funcional. En consecuencia, mi propuesta reside en entenderlo como un concepto teórico, cuyas condiciones de determinación vimos a lo largo de este apartado.
5.2. El lenguaje funcional en el estudio de las bases neurales del comportamiento
La atribución de la función biológica primaria al canto de llamada puede encontrarse en diversos trabajos neuroetológicos dedicados a indagar qué características de los cantos resultan atractivas para las hembras y cuáles son las bases neurales de la fonotaxis. Tales trabajos, de una forma u otra, utilizan un lenguaje funcional (note el lector mi énfasis en las citas) para destacar dicho efecto relevante del canto de llamada:
El canto del grillo más común es el canto de llamada, la cual es cantada por los machos para guiar a las hembras sexualmente receptivas hacia la madriguera del cantor (Bentley & Hoy, 1974, p. 34, mi énfasis).
En las ranas arbóreas y los grillos, por ejemplo, los machos producen sonidos que sirven para atraer hembras conespecíficas receptivas para el apareamiento (Doherty, 1985a, p. 279, mi énfasis).
El canto de llamada masculina en los grillos es importante para atraer hembras receptivas, conespecíficas, para el apareamiento (Doherty, 1985b, p. 787, mi énfasis).
Los grillos están entre esos insectos que producen sonidos y los usan para la formación de parejas y el apareamiento subsecuente, para el espaciamiento de los machos y para la defensa territorial. Los cantos de llamada atraen a las hembras listas para la copulación y los cantos de cortejo guían a las hembras a adoptar la posición correcta para el apareamiento (Huber, 1990a, p. 300).
El comportamiento de orientación por medio del sonido ejecutado por las hembras, denominado fonotaxis, tendría, a su vez, la función de hallar a un macho específico de su especie (cf. sección 4.4). De este modo, comienza la indagación de las bases neurales de la fonotaxis, cuyo lenguaje funcional analizaré a continuación.
El comportamiento fonotáctico de las hembras, cuya función es el hallazgo de un macho conespecífico (Schoneich y Hedwig, 2010), fue analizado como compuesto por dos tareas (cf. sección 4.4): la tarea de reconocimiento del canto conespecífico y la tarea de localización del macho emisor. La cuestión por la forma en la que dichas tareas son resueltas por el sistema nervioso de las hembras de grillo dio lugar a una serie de estudios neuroetológicos. Como vimos (sección 4.4.1) los estudios comportamentales intentaron establecer, en primer término, qué parámetros del canto de llamada son los que resultan atractivos para las hembras. En términos de Tinbergen (1951), podría decirse que estos estudios comportamentales intentaron aislar el estímulo señal que es relevante para la ejecución de la respuesta en cuestión. Tales estudios lograron determinar, con cierto grado de consenso científico, que la característica relevante de los cantos de llamada es el parámetro de la tasa de sílabas por segundo. Con este resultado, en segundo término, se ofrecieron diversos modelos con el fin de explicar cómo tales parámetros son procesados por los sistemas nerviosos de las hembras a los fines de ejecutar la respuesta fonotáctica, y se indagaron las estructuras y actividades de diversas neuronas que podrían constituir su implementación neural (cf. sección 4.4.2).
En este punto, la indagación sobre cómo las hembras resuelven la tarea de reconocimiento del canto conespecífico involucró el delineamiento de un modelo funcional (en un sentido que intentaré especificar) que explique el modo en el cual dicha tarea es resuelta, y la búsqueda del mecanismo neural subyacente que implementaría dicho modelo. El modelo funcional particular que hemos abordado (sección 4.4.2), el modelo de “autocorrelación”, apelaba a ciertas funciones, tales como “vía directa”, “vía de retardo”, etc., cuya organización era representada con el siguiente esquema:

Figura 11: modelo de “autocorrelación” propuesto para el reconocimiento del periodo de sílabas específico de la especie (Schöneich, Kostarakos y Hedwig, 2015).
La cuestión por el significado y el uso del lenguaje funcional en esta instancia de la investigación puede analizarse fructíferamente en términos de la propuesta ofrecida por el enfoque sistémico (cf. sección 3.2). Según este enfoque, las atribuciones funcionales sistémicas se determinan en el contexto de un análisis funcional adecuado de una disposición o capacidad explanandum ψ. En el contexto de esta investigación, el explanandum se identificaría con la capacidad de las hembras de grillo de hallar a un macho conespecífico, o con la manifestación de la disposición (el efecto) de dirigirse hacia un macho conespecífico en presencia de las condiciones precipitantes, constituidas por el canto de llamada (o la tasa de repetición de sílabas) específica de la especie (estímulo señal).
Dado este explanandum, el análisis funcional sistémico explicaría cómo las manifestaciones de la disposición (el hallazgo del macho conespecífico) son provocadas por las condiciones precipitantes (el canto de llamada conespecífico) ofreciendo un análisis de la capacidad explanandum en ciertas sub-disposiciones o sub-capacidades organizadas de una determinada manera (un modelo funcional). En este punto, un primer análisis funcional sistémico puede ser identificado cuando los científicos indican que, para el hallazgo de un macho conespecífico, el sistema nervioso de las hembras debe resolver dos tareas: la tarea de reconocimiento del canto y la tarea de localización del emisor (cf. sección 4.4). De este modo, el primer análisis quedaría establecido como sigue:
1. Primer análisis funcional sistémico:
Disposición o capacidad Ψ de S: Hallazgo de un macho conespecífico a partir del canto de llamada (Ψ) por parte de las hembras de grillo (S).
Análisis funcional de Ψ de S: {φ1; φ2}
En donde:
φ1: capacidad de S de reconocer el canto conespecífico, o disposición de S a responder (fonotácticamente) únicamente ante la tasa de repetición de sílabas específica de la especie.
φ2: capacidad de S de localizar al macho emisor, o disposición de S a dirigir su movimiento hacia la localización del macho a partir de su canto de llamada.
Como vimos (sección 4.4.2), el modelo de “autocorrelación” tiene como objetivo explicar el modo en el cual las hembras son capaces de reconocer el canto conespecífico (φ1). En este punto, este modelo toma como explanandum una capacidad establecida por el primer análisis funcional sistémico, y ofrece una descomposición de dicha capacidad en otras sub-capacidades, cuya manifestación ordenada sería equivalente a la manifestación de la capacidad φ1. De este modo, el segundo análisis funcional sistémico quedaría establecido como sigue:
2. Segundo análisis funcional sistémico:
Disposición o capacidad φ1 de S: capacidad de reconocer la tasa de repetición de sílabas específica de la especie, o disposición a responder (fonotácticamente) únicamente ante la tasa de repetición de sílabas específica de la especie.
Análisis funcional de φ1 de S: {α1; α2; α3; α4}
En donde:
α1: “vía directa” [auditory pathway]: capacidad de transmitir directamente (sin retardo temporal) el patrón de pulsos recibido hacia el “detector de coincidencias”.
α2: “vía de retardo” [delay line]: capacidad de transmitir el patrón de pulsos recibido hacia el “detector de coincidencias” con un retardo temporal correspondiente al periodo de pulsos específico de la especie.
α3: “detector de coincidencias” [coincidence detector]: capacidad de estimular el “detector de características” cuando los estímulos de la “vía directa” y la “vía de retardo” recibidos coinciden temporalmente.
α4: “detector de características” [feature detector]: capacidad de responder únicamente al patrón de pulsos específico de la especie.
En este punto, los investigadores procedieron a indagar el modo en el cual este modelo podría estar implementado en el sistema nervioso de las hembras. Con este objetivo, indagaron los patrones de actividad de ciertas neuronas del protocerebro anterior de las hembras, comparando sus respuestas ante distintos patrones de sonido presentados, incluyendo en la presentación el patrón de pulsos específico de la especie. El resultado de esta indagación permitió a los investigadores atribuir a las neuronas examinadas y a sus patrones de respuesta la implementación del modelo funcional presentado anteriormente. Esto es: esta indagación permitió explicar las capacidades {α1; α2; α3; α4} del modelo funcional precedente a partir de la actividad de ciertas neuronas {AN1; LN2, LN5; LN3; LN4} y de su modo de organización. De este modo, la explicación de cómo estas capacidades son instanciadas apela, nuevamente, a otras capacidades, pero, esta vez, a capacidades de partes del sistema y a su modo de organización. Procediendo de esta manera, el análisis funcional sistémico logra identificarse con un análisis funcional mecanicista, en tanto logra mostrar cómo las capacidades analizantes propuestas son instanciadas por las actividades de ciertas entidades y su modo de organización, esto es, en tanto logra mostrar cómo las capacidades analizantes propuestas y su modo de organización son instanciadas en un mecanismo. En este caso, las capacidades analizantes propuestas por el análisis funcional sistémico (2) se instanciarían de la siguiente manera:
3. Análisis funcional mecanicista:
α1: “vía directa” [auditory pathway]: se instancia en la actividad de disparo AN1, que transmite los patrones de pulsos recibidos, mediante inputs excitatorios, hacia LN2 y LN3.
α2: “vía de retardo” [delay line]: se instancia en la actividad de disparo de LN2 y LN5. Ante la recepción de inputs excitatorios por parte de AN1, LN2 inhibe a LN4 y a LN5. Ante esta inhibición, LN5 excita LN3 con un retardo de 40mseg (PIR).
α3: “detector de coincidencias” [coincidence detector]: se instancia en la actividad de disparo de LN3, que causa una despolarización supra-umbral en LN4 cuando los inputs excitatorios recibidos de AN1 y LN5 coinciden temporalmente.
α4: “detector de características” [feature detector]: se instancia en la actividad de disparo de LN4, que supera la inhibición de LN2 y dispara potenciales de acción cuando recibe el input excitatorio suficiente para dicha superación por parte de LN3.
Dado que para el enfoque sistémico las atribuciones funcionales se identifican con la identificación de sub-capacidades analizantes (de sistemas o de partes de sistemas) que explican la implementación de una capacidad explanandum, en este contexto las atribuciones funcionales estarían determinadas por cada análisis funcional realizado en cada instancia de esta investigación. Así, dentro del primer análisis funcional sistémico (1) de la capacidad de las hembras de hallar un macho conespecífico, se habrían determinado dos funciones: la función de localización y la función de reconocimiento. Luego, dentro del segundo análisis funcional (2) de la capacidad de reconocimiento, se habrían determinado cuatro funciones: la función vía directa, la función vía de retardo, la función detector de coincidencias y la función detector de características. Por último, dentro del análisis funcional mecanicista de estas funciones, se habrían determinado las funciones de las cinco neuronas que implementan las funciones del segundo análisis funcional. En este punto, las actividades de tales neuronas detalladas en (3) constituirían sus “roles funcionales mecanicistas” (Craver, 2001), esto es, aquellas propiedades o actividades en virtud de las cuales contribuyen a la capacidad de reconocimiento del mecanismo del que participan.
Si bien el enfoque sistémico permite dar cuenta de la manera en la que los neuroetólogos utilizaron el lenguaje funcional al indagar las bases neurales del comportamiento fonotáctico de las hembras, debemos recordar que, según este enfoque, las capacidades explanandum a las cuales se aplica el análisis funcional se encuentran débilmente restringidas por criterios teóricos y pragmáticos (cf. sección 3.2). La escasa limitación de las capacidades explanandum a las cuales se aplica el análisis llevó a muchos autores a formular distintas objeciones y contraejemplos que destacan cierta promiscuidad en las adscripciones funcionales que permitiría la teoría. Sin embargo, teniendo en cuenta nuestro análisis de la sección anterior, referida a la atribución funcional del comportamiento de llamada de los machos y la fonotaxis de las hembras, parece posible decir algo más respecto de la selección de la capacidad explanandum a la que se aplicó fructíferamente el análisis funcional.
En primer lugar, en cuanto a los criterios teóricos utilizados para seleccionar dicho explanandum, y según lo visto en el apartado anterior, la capacidad fonotáctica de las hembras y su función (el hallazgo de machos conespecíficos) alcanzó relevancia explicativa en tanto daba cuenta de cómo los grillos logran formar parejas reproductivas. El canto de llamada del macho, junto con su función de atraer hembras conespecíficas, es la contracara del mismo fenómeno: la formación de parejas reproductivas, que es uno de los eventos que constituyen la “secuencia reproductiva” (Alexander, 1997). Si bien es posible dar cuenta de estas atribuciones funcionales, en parte, apelando a un análisis funcional “bottom-up” (cf. sección 5.1.2), como vimos, ninguno de los criterios utilizados por los biólogos constituía una condición necesaria y suficiente para la determinación de la función de tales comportamientos. Como sugerí antes, esta característica de las atribuciones funcionales biológicas no es problemática si uno entiende que el concepto de función es un concepto teórico. En este punto, sugiero, la promiscuidad a la que puede dar lugar la aplicación del análisis funcional se evita, en este contexto, aplicando el análisis a capacidades a las cuales se las considera, teóricamente, funciones biológicas. Pero esto tiene una consecuencia importante en lo que respecta a la evaluación del enfoque sistémico. Este enfoque resultó adecuado para dar cuenta de algunos aspectos de la práctica científica, pero no logra dar cuenta, al menos de manera completa, de la atribución de la función biológica. Y es esta atribución la que permite acotar la promiscuidad del análisis funcional, volviendo a este tipo de análisis relevante y explicativo.
En segundo lugar, en cuanto a los criterios pragmáticos utilizados para seleccionar la capacidad explanandum, parece posible especificar qué tipo de criterios pragmáticos utilizan los neuroetólogos recordando el rol restrictivo de la teoría de la selección natural que explicité en la sección 5.1. ¿Por qué a los neuretólogos les interesa explicar ciertas capacidades por sobre otras? ¿Por qué eligen explicar capacidades que consideran funciones biológicas? Si bien ofrecer una respuesta a estas preguntas puede ser considerado un asunto de especulación psicológica, una respuesta posible consiste en afirmar que se eligen tales capacidades en tanto son consistentes con, y explicables por, la teoría de la selección natural darwiniana. Si bien la teoría de la selección natural no determina el tipo de funciones biológicas que cumplen los rasgos (cf. sección 5.1), ofrece un marco teórico en el cual ciertas capacidades tienen mayor relevancia que otras. Continuaré con estas reflexiones en el apartado siguiente dedicado a las conclusiones.
5.3. Conclusiones parciales
A lo largo de este capítulo ofrecí un análisis del uso del concepto de función en la práctica de la neuroetología en lo que refiere al caso de la investigación de la comunicación acústica del grillo de campo. Al mismo tiempo, contrasté la adecuación de los diversos enfoques presentados en el capítulo 3 para dar cuenta del uso del lenguaje funcional en este caso particular. Los resultados pueden ser resumidos en una serie de conclusiones.
En primer lugar, vimos que las atribuciones funcionales eran utilizadas por Alexander como parte de la evidencia para reconstruir la historia evolutiva del rasgo e inferir la acción de la selección natural en el pasado. Según Alexander, el origen de las distintas funciones que cumplen los cantos (de cortejo, de llamada, de agresión) habría dependido de ciertos cambios en la estructura del canto “original” (de cortejo) en los ancestros de los gríllidos y los tetigonideos, y de ciertas ventajas que habrían reportado dichos cambios para el cumplimiento de nuevas funciones (la atracción de hembras a distancia, el alejamiento de machos). Dado que las atribuciones funcionales constituyen en este caso parte de la evidencia para la reconstrucción de dicha historia, y dado que el origen de dichas funciones es aquello que es explicado por la teoría de la selección natural, uno de los resultados de este análisis consiste en que el concepto de función, en contra del enfoque etiológico y de selección no histórica, es (y debe ser tratado como) un concepto independiente de la teoría de la selección natural. Si la teoría de la selección natural determinara aquello que constituye su evidencia y su explanandum, perdería explicatividad en tanto se tornaría circular. Por lo tanto, siendo evidencia y explanandum de la teoría, el concepto de función no puede ser parte de ella. Sin embargo, como sugerí antes, la teoría de la selección natural podría tener un rol heurístico en las atribuciones funcionales, en tanto restringiría el tipo de objetivos que los organismos pueden perseguir (rol restrictivo), pero sin cumplir un rol determinante a la manera que sugiere el enfoque etiológico y el enfoque de selección no histórica (en su versión etiológica). De este modo, en contra de estos enfoques, el resultado de esta indagación sugiere que la teoría de la selección natural es relevante para la atribución de funciones, en tanto puede poseer un rol heurístico, pero dicha atribución no debe ser confundida con la determinación del origen histórico del rasgo. Esto nos llevó a indagar el tipo de evidencia que utilizaron los científicos para realizar dicha atribución funcional, lo cual nos lleva al siguiente punto.
En segundo lugar, lo largo de la sección 5.1.2 expuse diversos criterios que fueron utilizados en el caso de la atribución funcional del canto de llamada. En el examen de dicha evidencia vimos cómo el enfoque sistémico y el enfoque de valor de supervivencia podían dar cuenta de la utilización de algunos criterios utilizados por los científicos para determinar dicha atribución funcional. Además, la formulación de razonamientos contrafácticos del tipo sugerido por Canfield (1964) y por Bigelow y Pargetter (1987) (cf. sección 3.3) sobre las posibilidades de supervivencia y/o reproducción que habrían resultado si el efecto funcional no se diera, parecía proponer no sólo un criterio adecuado para la atribución funcional, sino un fin último de los organismos consistente con la teoría de la selección natural en su rol heurístico. Sin embargo, la sugerencia de Horan y del enfoque de selección no histórica (en su versión de valor de supervivencia) de que tales atribuciones se determinan realizando mediciones de fitness resultó inadecuada, en tanto tales mediciones se llevan adelante, en la práctica, presuponiendo cierto concepto de función biológica en el sentido aquí esbozado. Así, como vimos en este caso, los estudios que apuntaron a establecer la contribución al fitness de un determinado comportamiento presupusieron algún sentido de función determinado previamente. La contribución del comportamiento de llamada al “éxito en el apareamiento” (lo que era considerado un proxy del fitness) era comparado con la contribución del comportamiento “satélite” bajo ciertas condiciones ambientales (la densidad de población). Ambos comportamientos eran considerados “estrategias reproductivas”, lo cual involucra considerarlos comportamientos reproductivos alternativos, o comportamientos que cumplen una determinada función en la consecución del objetivo de la reproducción. Y como vimos, si el desempeño de dicha función contribuye al fitness o no era dependiente de ciertas condiciones ambientales, lo cual no parece ser cierto de este sentido de función. Así, el comportamiento de llamada, o el comportamiento satélite, son estrategias reproductivas aunque no contribuyan al fitness bajo ciertas condiciones, e incluso son estrategias reproductivas de manera previa a conocer bajo qué condiciones realizan tal contribución.
Como vimos, además, ninguno de los criterios utilizados por los científicos para la determinación de dicho concepto de función, ni todos en conjunto, constituían propiamente condiciones necesarias y suficientes para la aplicación del concepto, esto es, una definición del concepto. Por el contrario, dichos criterios sugerían que el concepto de función, en este sentido, podría ser un concepto teórico cuyos criterios de determinación constituyen “síntomas” para su aplicación. Por lo tanto, un segundo resultado de este análisis es prospectivo: podría ser fructífero entender este concepto de función (que aquí denomino “función biológica”) como un concepto teórico e intentar dilucidar la teoría subyacente, como ya se realizó en algunas ocasiones (Ginnobili, 2009, 2011).
Por último, a lo largo de 5.3 vimos que el estudio de las bases neurales de la fonotaxis implicó, en el contexto de una serie de análisis funcionales, la determinación de diversas funciones sistémicas. El análisis funcional procedió, en primer lugar, descomponiendo o analizando la “función biológica” de la fonotaxis determinada previamente (el efecto de hallar un macho conespecífico a partir de su canto de llamada) en dos funciones o “tareas”: la tarea de reconocimiento de la llamada conespecífica, y la tarea de localización de su emisor. En el análisis funcional de la tarea de reconocimiento, se determinaron, a su vez, un conjunto de funciones que darían cuenta de cómo la función o capacidad de reconocimiento de las hembras puede manifestarse a partir de sus condiciones precipitantes (el patrón de pulsos específico de la especie): la vía directa, la vía de retardo, el detector de coincidencias y el detector de características. Dicha determinación culminó en la atribución de roles funcionales mecanicistas a ciertas neuronas del sistema nervioso de las hembras (AN1, LN2, LN3-3*, LN4, LN5) que implementarían dichas funciones sistémicas en tanto poseen cierta estructura y manifiestan ciertas actividades. Además, vimos cómo el análisis funcional evita, en este caso, la carga de promiscuidad tomando como capacidad explanandum dicha función biológica (¿cómo las hembras logran hallar a los machos conespecíficos a partir de su canto?). Por otra parte, dado que el análisis funcional explica, justamente, cómo las manifestaciones de la capacidad son provocadas por las condiciones precipitantes (sección 3.2), parece necesario primero conocer cuáles son tales condiciones o, como fue en este caso de estudio, cuál es el estímulo señal o releaser de la respuesta fonotáctica de las hembras. Como vimos, la indagación sobre el estímulo señal al que responden las hembras fue previa al estudio de las bases neurales de su procesamiento, y el conocimiento de dicho estímulo fue necesario en orden de correlacionar la actividad de ciertas neuronas con los patrones de sonido presentados. Por lo que, en este caso, creo que puede apreciarse el modo en que la atribución funcional biológica operó evitando la promiscuidad de las atribuciones funcionales sistémicas: permitiendo hallar no sólo la capacidad relevante, sino el estímulo relevante ante el cual dicha capacidad se manifiesta. Y dicho estímulo relevante es, precisamente, el canto de llamada, cuya atribución funcional analicé previamente.
- Agradezco a Ariel Roffé por el diseño del esquema.↵
- Véase Searcy & Anderson (1986) para una recopilación y discusión de las distintas hipótesis posibles acerca de la evolución de los cantos de llamada y las preferencias de las hembras.↵
- Hunt & Hodgson (2010) consideran que el éxito en el apareamiento es un proxy del fitness utilizado frecuentemente en ecología del comportamiento, por lo que en este trabajo no cuestionaré dicha asunción. El lector puede encontrar en dicho trabajo una presentación de las distintas formas en las que puede medirse el fitness y las dificultades que acarrea cada método de medición.↵

